
Revista Científica UDO Agrícola Volumen 9.
Número 4. Año 2009. Páginas: 802-806
Influencia del plaguicida cloropirifos
sobre la fotosíntesis, transpiración y conductancia estomática en yerba mate (Ilex paraguariensis A. St.-Hil.)
Effect of chloropyrifos on photosynthesis, transpiration and stomatal conductance of yerba mate (Ilex paraguariensis A.St.-Hil.).
Alicia Emilia CASTILLO ![]() , Pedro Alfonzo SANSBERRO y Claudia Verónica LUNA
, Pedro Alfonzo SANSBERRO y Claudia Verónica LUNA
Facultad de Ciencias Agrarias. Universidad
Nacional del Nordeste. Sargento Cabral 2131. Corrientes. Argentina. E-mails:
aecast@yahoo.com.ar y castillo@agr.unne.edu.ar
![]() Autor para
correspondencia
Autor para
correspondencia
|
Recibido:
04/08/2009 |
Fin de
primer arbitraje: 29/09/2009 |
|
Primera
revisión recibida: 15/11/2009 |
Aceptado: 12/12/2009 |
RESUMEN
Este trabajo tuvo como objetivo observar los efectos
del pesticida cloropirifos sobre la conductancia
estomática, fotosíntesis y transpiración
en yerba mate (Ilex paraguariensis A.
St.-Hil.).
Se realizó un ensayo en cámara con condiciones controladas y plantas de yerba
mate, una por maceta, a las que
se les aplicaron tres dosis de cloropirifos (50, 100 y 150 ppm) y un testigo sin
pesticida. Uno, 7 y 21 días después de la aplicación se evaluaron la
fotosíntesis, transpiración y conductancia estomática. Ninguna de las variables
evaluadas fue afectada con las dosis aplicadas. Sin embargo, con la edad de las
plantas incrementaron significativamente la conductancia estomática (300%), la
fotosíntesis (116%) y la transpiración (285%) entre los 7 y 21 días.
Palabras clave: Cloropirifos,
fisiología vegetal, tasa fotosintética, yerba mate.
ABSTRACT
The objective was to evaluate the effect of chlorpyrifos
on stomatal conductance, photosynthesis and
transpiration on yerba mate (Ilex paraguariensis A. St.-Hil.) plants. The experiment was conducted in a chamber under controlled conditions.
The plants (one/pot) were sprayed with three doses (50, 100 and 150 ppm of chlorpyrifos) and a non pesticide-sprayed
control. Stomatal conductance, photosynthesis and
transpiration were evaluated at 1, 7 and 21 days after pesticide application.
The mentioned traits were not affected by the pesticide application doses.
However, significative differences were found as the
plants were growing and traits increased between 7 and 21 days in 300% for stomatal conductance, 116% for photosynthesis and 285% for
transpiration.
Key words: Chlorpyrifos, vegetal physiology,
photosynthetic rate, Ilex paraguariensis.
Introducción
El uso de pesticidas
es a menudo necesario en la producción vegetal, para prevenir daños de
insectos en los cultivos. Sin embargo, su uso puede llegar a niveles de
toxicidad y afectar negativamente el
crecimiento y desarrollo vegetal. El impacto de los insecticidas en los procesos de intercambio
gaseoso en la planta es poco entendido,
aunque su uso extensivo se ha sugerido como
limitativo para la productividad de las cosechas. Si un insecticida
altera la fisiología vegetal, la cosecha puede ser afectada (Qiu et al.,
2002). Algunos estudios han demostrado efecto significativo de los insecticidas
sobre la fisiología de la alfalfa (Medicago sativa
L.) y la soja (Glycine max (L.) Merrill) (Haile
et al., 1999).
Los insecticidas aplicados en el follaje
pueden afectar la fotosíntesis, por el
cierre parcial de los estomas. A la vez, estos efectos
pueden retrasar o disminuir la calidad de la producción. Es el caso de la
actividad fotosintética de la lechuga que disminuyó en forma no permanente con
la aplicación de algunos insecticidas, aunque su biomasa seca final se mantuvo
sin cambio (Haile et
al., 2000). Por lo tanto, parece necesario entender los efectos de los
insecticidas, usados más frecuentemente, en la fisiología vegetal y regular
esos efectos (Untiedt y Blanke, 2004).
Se han realizado distintos estudios
referidos a evaluar el comportamiento de los diferentes fenómenos fisiológicos,
luego de la aplicación de pesticidas. Por ejemplo, los índices fotosintéticos
de lechuga fueron reducidos sensiblemente
por algunos insecticidas a partir de las cuatro horas de una pulverización. Sin embargo, la
reducción en fotosíntesis de la lechuga por dichas aplicaciones era solamente temporal, de tal forma que no hubo una alteración
perceptible del peso seco de la planta (Haile et al., 2000).
Entre los pesticidas más usados se
encuentran los organofosforados, como el cloropirifos.
El efecto que producen estos pesticidas en los seres vivos es la inhibición de la acetilcolinesterasa, una
enzima que impide la acción de la acetilcolina por catálisis de su hidrólisis,
con lo que se interrumpe el funcionamiento normal de la transmisión de impulsos
nerviosos en animales. Sus residuos pueden persistir en las plantas de
La
yerba mate (Ilex paraguariensis A. St.-Hil.) es un
árbol o un arbusto cuyas hojas secas y trituradas se utilizan en regiones de
Sudamérica para preparar una bebida tradicional que data de las épocas
pre-colombinas. La República Argentina, favorecida por sus condiciones
agroclimáticas, es el principal productor mundial, la producción nacional de
yerba mate canchada (seca) en el año 2008 fue de 235,1 millones de kilogramos
concentrándose su cultivo en las provincias de Misiones (88,9%) y Corrientes
(11,1%) donde se registran 202.736,33 ha plantadas con yerba mate (INYM,
2009). Por su parte, Brasil, caracterizado por un
sistema de explotación extractivo de los montes nativos, posee 85.000 ha,
mientras que en Paraguay ésta alcanza a 35.000 ha (Gortari, 2007).
Por el volumen de producción e
importancia económica que representa, es necesario mantener un nivel de sanidad
optimo, básicamente de las hojas, que seria la
materia prima, ya que el cultivo cuenta con algunas plagas importantes tales
como, el “Rulo” o Psílido de la yerba mate (Gyropsylla spegazziniana
Lizer), el taladro grande o “tigre de la yerba mate”
(Hedyphates betulinus Klug), el marandová de la yerba
mate u oruga rabuda, (Perigonia lusca F.) y
ácaros, que en su mayoría son controlados con productos organofosforados (Burtnik, 2003; CEUTA, 2006).
El objetivo de este trabajo fue evaluar
el efecto de la aplicación de cloropirifos sobre la
conductancia estomática, fotosíntesis y transpiración en yerba mate, en
condiciones controladas de crecimiento.
MATERIALES
Y MÉTODOS
Experimento
Fue establecido en macetas en
cámara climatizada bajo condiciones controladas con una planta por maceta y una
maceta por repetición. Los tratamientos fueron: Control (rociado con agua) (T0)
y tres dosis de cloropirifos: (T1) 50 ppm, (T2) 100 ppm, y
(T3) 150 ppm. El insecticida fue aplicado solamente al inicio de la
experiencia, teniendo en cuenta que la vida media del producto generalmente
está entre 60 y 120 días. Se cuantificó la tasa de fotosíntesis, conductancia
estomática y transpiración a los 1, 7 y 21 días después de la aplicación del
producto.
Pesticida
Se usó el insecticida de nombre comercial
TauroL. O, en forma de emulsión con 48% de cloropirifos.
Material
vegetal y condiciones del ensayo
Las plantas de yerba mate
pertenecían al clon SI-49 (Las Marías Co.), de
aproximadamente 4 años de edad, pertenecientes al vivero del establecimiento.
Las mismas se transplantaron a macetas de polietileno
de 4 L, con suelo rojo del orden ultisol (Gob. Virasoro, Corrientes). El suelo se mantuvo a capacidad de
campo y el riego fue diario. Las condiciones dentro de la cámara climatizada
fueron: temperaturas entre 27 ± 1 y 22 ± 2 ºC día/noche, fotoperíodo de 14
h, densidad de flujo fotosintético del fotón
400 µmol.m-2.s-1
a partir de lámparas fluorescentes y
humedad relativa entre 50 y 55%.
Intercambio
gaseoso foliar y medición de conductancia estomática
Las tasas de intercambio gaseoso foliar y la conductancia estomática (gs) fueron
evaluadas en hojas completamente maduras
y enteras, localizadas en el 6to nudo a partir del ápice y en horas
del mediodía, con un analizador de gases en el infrarrojo, con el
sistema de intercambio abierto (Li-Cor LI-6400). La fuente de luz LI-6400 fue utilizada para controlar las densidades
de flujo fotosintéticas del fotón (PPFD) a
1800 mmol m-2 s-1 . La corriente de aire de entrada fue mantenida a 350 mmol CO2·mol-1 con un sistema
de mezcla controlado de CO2,
suministrado por el LI-6400. Las hojas,
la cubeta, y temperatura del aire fueron
medidas con termocuplas.
La temperatura de la cámara de medición y el tejido de la hoja fue mantenida a
27 ºC con un módulo Peltier
controlado y colocado sobre la cubeta. Esta temperatura fue equivalente a la
media del ambiente dentro de la cámara de ambiente controlado. El déficit de la presión de vapor de hoja a aire en la cámara
fue mantenido aproximadamente a 1,6 KPa.
Análisis
estadístico
Se utilizó un diseño estadístico de bloques completos al azar, con tres
repeticiones y una maceta con una planta como unidad experimental. Se realizó
análisis de varianza y se utilizó la
prueba de Duncan de comparación múltiple
para evaluar diferencias entre las medias de los tratamientos (p< 0,05).
RESULTADOS
Los resultados del presente estudio podrían
representar la primera aportación de los efectos fisiológicos del cloropirifos en yerba mate. Actualmente existe información
equivalente con dimetoato (Schmalko
et al. ,2002). La fotosíntesis el
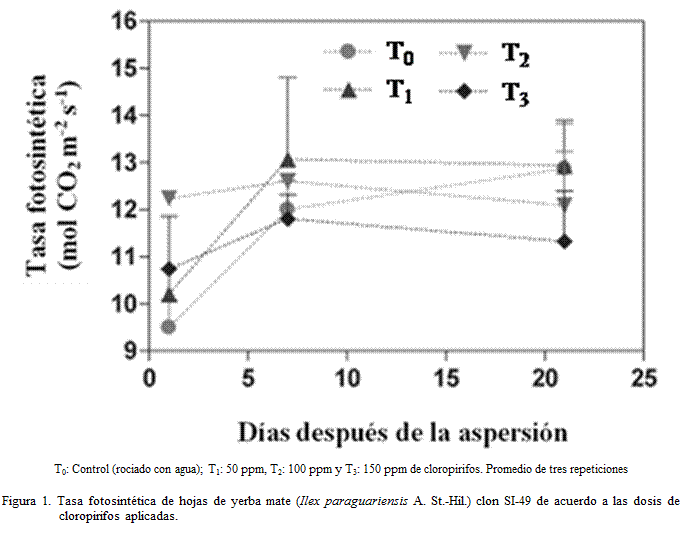
primer día fue mayor en los tratamientos con el pesticida respecto al control.
En los siguientes días no se observó una tendencia única; aunque el menor valor se observó en el tratamiento
con la dosis mayor (Figura 1). Comportamiento similar fue observado en
berenjena (Solanum melongena L.)
por Yu et al.
(2006).
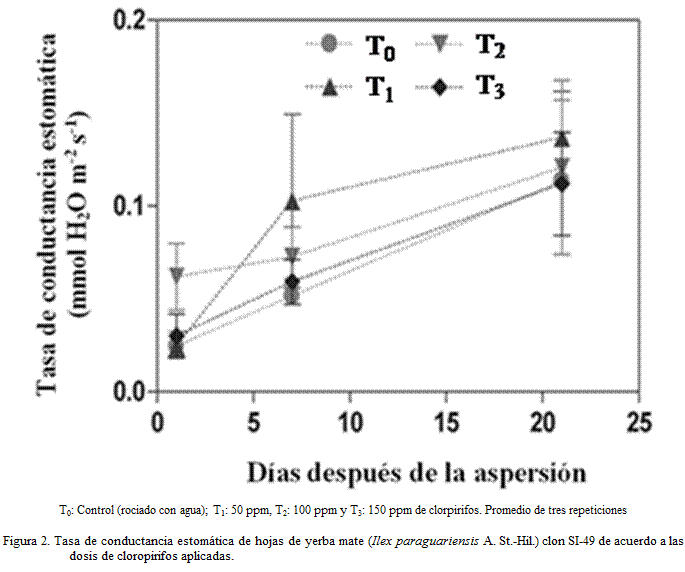
La conductancia estomática en el T0 presentó el valor menor, el T3 presentó
niveles similares, mientras que el T1 tuvo un incremento sostenido
(Figura 2).
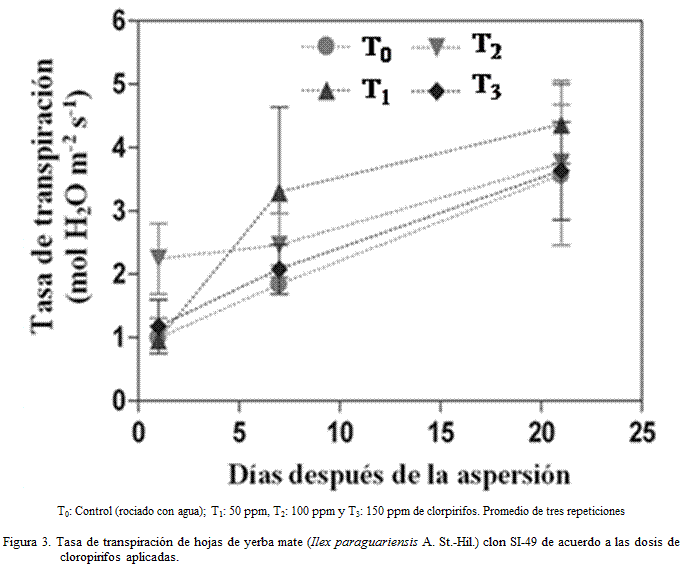
El T0 mantuvo la menor transpiración durante
los 21 días de evaluación, el T3 mantuvo valores similares a T0.;
mientras que el T1 tuvo un crecimiento sostenido (Figura 3).
El análisis estadístico indicó que las
medias totales de fotosíntesis, transpiración y conductancia estomática fueron
estadísticamente similares entre las dosis de clorpirifos
aplicadas (Cuadro 1) y los valores medios incrementaron significativamente a
través del tiempo en todos los tratamientos; aunque, el nivel menor correspondió al tratamiento con
la mayor dosis aplicada. Diferencias altamente significativas fueron observadas
en la fotosíntesis, transpiración y conductancia estomática entre el primer día
y 21 días posteriores a la aplicación del plaguicida (Cuadro 2) en los
tratamientos con y sin insecticida.
|
Cuadro 1. Niveles medios totales de tasa
fotosintética, conductancia estomática y transpiración de hojas de
yerba mate (Ilex paraguariensis A.
St.-Hil.) clon
SI-49 de acuerdo a las dosis de
cloropirifos aplicadas.* |
||||||
|
Dosis |
Tasa fotosintética (mol CO2 m-2 s-1) |
Tasa de conductancia estomática
(mmol H2O m-2 s-1) |
Tasa de transpiración (mol H2O m-2 s-1) |
|||
|
T3 |
11,29 |
A |
0,06 |
A |
2,13 |
A |
|
T0 |
11,46 |
A |
0,07 |
A |
2,29 |
A |
|
T1 |
12,07 |
A |
0,09 |
A |
2,82 |
A |
|
T2 |
12,31 |
A |
0,09 |
A |
2,88 |
A |
|
T0: Control (rociado con agua); T1:
50 ppm, T2: 100 ppm y T3: 150 ppm de cloropirifos. Promedio de tres repeticiones *
Letras iguales no difieren estadísticamente (p< 0,05). |
||||||
|
Cuadro 2.
Niveles medios totales de tasa fotosintética, conductancia estomática y
transpiración de hojas de yerba mate (Ilex paraguariensis A. St.-Hil.) clon SI-49 de acuerdo a los días transcurridos después de la
aplicación de cloropirifos.* |
||||||
|
Días ** |
Tasa fotosintética (mol CO2 m-2 s-1) |
Tasa de conductancia
estomática (mmol H2O m-2 s-1) |
Tasa de transpiración (mol H2O m-2 s-1) |
|||
|
1 |
10,67 |
A |
0,04 |
A |
1,34 |
A |
|
7 |
12,30 |
B |
0,07 |
A |
2,42 |
B |
|
21 |
12,37 |
B |
0,12 |
B |
3,83 |
C |
|
* Letras
iguales no difieren estadísticamente (p< 0,05). ** Días despúes de la aspersión de cloropirifos |
||||||
DISCUSIÓN
Los
resultados obtenidos pueden indicar que las concentraciones empleadas de
cloropirifos no afectan las tasas fotosintética y trasnpiratoria y la conductancia estomática de yerba mate (Ilex paraguariensis A.
St.-Hil.) clon
SI-49 durante los primeros 21 días. Estos
resultados son parcialmente parecidos a los encontrados por otros autores en especies, como G. max. Así,
en soja se observó que los índices de la conductancia estomática y
transpiración fueron significativamente similares entre los tratamientos con
insecticida (Haile et al., 1999). En contraste, otros autores encontraron que esta especie
disminuye su tasa fotosintética en los
primeros días posteriores a la aplicación, respecto al testigo (Abdel-Reheem et al.,
1991; Li et al., 2008).
Para la alfalfa los niveles
fotosintéticos encontrados no presentaron
diferencias ante algunos insecticidas probados (piretroides, permetrina, cloropirifos, carbofuran, carbaril y espinosad) estos
insecticidas son químicamente disímiles)
en distintas etapas del crecimiento
o reproductivas (Haile et al., 1999).
La
aplicación de cloropirifos en las dosis aplicadas y
su posible efecto sobre la fotosíntesis
se han investigado también en pepino (Cucumis sativus L.). En esa especie se observó que la
inhibición de la fotosíntesis fue acompañada por la disminución en la
conductancia estomática (Xia et al., 2006). Sin
embargo, el resultado significativo del
efecto de los pesticidas se ha podido observar después de algunos días de la
aplicación. La tasa fotosintética de lechuga en almácigo fue reducida por algunos insecticidas (endosulfan,
metomil, acefato) a partir
de las 4 h y hasta los 2 días después de la aplicación. Dicha reducción sólo fue transitoria, debido a
que la lechuga recuperó los niveles de fotosíntesis considerados normales a
partir de los 5 días de que la aplicación fuera hecha. No se encontraron
diferencias en el rendimiento de su biomasa seca
(Haile et al.,
2000).
CONCLUSIÓN
El cloropirifos
en dosis entre 50 y 150 ppm no afectó la fisiología de la yerba mate. El
incremento de las tasas fotosintética y transpiratoria y la conductancia estomática aumentaron con el tiempo de la aplicación y no se puede
señalar que sea resultado de la degradación del pesticida ya que el testigo
tuvo el mismo comportamiento. Se sugiere
realizar nuevas evaluaciones con otros niveles de pesticida y tiempo más
prolongado.
LITERATURA CITADA
Abdel Reheem, S.; M. H. Belal and G. Gupta.
1991.
Photosynthesis inhibition of soybean leaves by insecticides .
Environ. Poll. 74:
245-250.
Burtnik, O. 2003. Manual del pequeño productor yerbatero correntino. INTA-Secretaria de Extension Rural Santo Tomé, Corrientes, Argentina. 30 p.
Centro Uruguayo de Tecnologías
Apropiadas (CEUTA). 2006. Agrotóxicos en Uruguay:
miradas desde los afectados. Montevideo, Uruguay. 58 p.
Gortari, J. 2007. El Instituto
Nacional de la Yerba Mate (INYM) como dispositivo político de economía social:
mediación intrasectorial en la distribución del
ingreso, empoderamiento del sector productivo y desarrollo local en la región
yerbatera. IADE. Buenos Aires. Realidad Económica 232: 21-31.
Haile F. J.; R. K. D. Peterson and L. G. Higley. 1999. Gas exchange responses of alfalfa and soybean
treated with insecticides. J. Econ. Entomol. 92: 954-959.
Instituto
Nacional de la Yerba Mate (INYM). 2009. Estadísticas de producción.
http://www.inym.org.ar/. Consultado el 12 de diciembre de 2009.
Li, W. J.; Z. K. Gao![]() , M. Wang, Y. P. Cao and Z. J. Xue. 2008. Toxicological effects of omethoate
on leaf photosystem II of cole. Chin. J. Appli.
Ecol. 19: 2317-2321.
, M. Wang, Y. P. Cao and Z. J. Xue. 2008. Toxicological effects of omethoate
on leaf photosystem II of cole. Chin. J. Appli.
Ecol. 19: 2317-2321.
Schmalko, M. E.; L. A. Ramallo, D. Ferreira and L. D. Belingheri. 2002. Dimethoate degradation in plants and during processing of
Yerba Mate Leaves. Braz. Arch. Biol. Tech.
45: 419-422.
Untiedt,
R. and M. M. Blanke. 2004. Effects of fungicide
and insecticide mixtures on apple tree canopy photosynthesis, dark respiration
and carbon economy. Crop Protect. 23: 1001-1006.
Untiedt,
R. and M. M. Blanke. 2004. Effects of fungicide
and insecticide mixtures on apple tree canopy photosynthesis, dark respiration
and carbon economy. Crop Protect. 23
(10): 1001-1006.
Página diseñada por Prof. Jesús Rafael
Méndez Natera
TABLA
DE CONTENIDO DE LA REVISTA CIENTÍFICA UDO AGRÍCOLA


