
Revista Científica UDO Agrícola Volumen 8.
Número 1. Año 2008. Páginas: 52-60
Efecto de la intensidad de luz sobre el
crecimiento del corocillo (Cyperus rotundus
L.)
Effect
of light intensity on growth of purple nutsedge (Cyperus
rotundus L.)
Nohelia M. RODRÍGUEZ R.![]() 1 y José
Vicente LAZO2
1 y José
Vicente LAZO2
1Insituto
Nacional de Investigaciones Agrícolas (INIA). CIAE Sucre-Nueva Esparta. Campo
Experimental Irapa, estado Sucre, Venezuela y 2Universidad
Central de Venezuela. Facultad de Agronomía. Instituto de Botánica Agrícola
Maracay Edo Aragua. E-mail: nrodriguez@inia.gob.ve ![]() Autor para
correspondencia
Autor para
correspondencia
|
Recibido: 09/06/2008 |
Fin
de primer arbitraje: 24/06/2008 |
Primera
revisión recibida: 05/08/2008 |
|
Fin
de segundo arbitraje: 25/08/2008 |
Segunda revisión
recibida: 06/09/2008 |
Aceptado: 10/09/2008 |
RESUMEN
Se determinó el efecto de la intensidad de la luz
sobre el crecimiento del corocillo, Cyperus rotundus
(L.). Los experimentos se realizaron en umbráculo y en el laboratorio, con suelo arenoso-arcillosos y los bulbos germinados bajo
oscuridad, con 3 hojas. Se utilizó un diseño estadístico completamente
aleatorizado, con tres repeticiones y tres tratamientos de intensidad de luz
(100, 50 y 25%). La temperatura media en el umbráculo fue de 28 °C (20-35 °C),
la humedad relativa media fue 60% (20-100%) y la radiación solar varió de 250 a
300 g.cal.cm-2.día-1. Se realizaron cinco muestreos cada
siete días de las partes aérea y subterránea de las plantas. Después de 21 días
de crecimiento, se observó un mayor número de hojas y una menor área foliar con
aumentos de la intensidad de la luz. Después de 28 y 35 días, el número de
hojas, área foliar y la longitud de la raíz disminuyeron, pero la masa de los
bulbos se incrementó. La floración tuvo lugar en la quinta semana. Después de
los 65 días en el 100% de tratamiento con luz, el número de hojas fue mayor,
mientras que el área foliar fue menor. La masa seca de la inflorescencia fue
menor en 25% de luz. Se concluye que la luz es importante en el crecimiento del
corocillo, debido a que una mayor intensidad de luz
promueve la floración y el número de hojas y mayor masa seca de las raíces,
bulbos y total. Para el control del corocillo, se
recomienda la aplicación de un sistema integral de sombra en combinación con un
herbicida y el control biológico.
Palabras clave: Cormos, corocillo, intensidad, luz, hojas.
ABSTRACT
The effect of light
intensity upon growth of purple nutsedge, Cyperus rotundus (L.) was determined. The experiments were made in shelter and laboratory, using
loamy-sandy soil and corms germinated under dark, with 3 leaves. A complete
randomized statistical design was used, with three replications and three light
intensity treatments (100, 50 and 25%). Average temperature in the shelter was
28 °C (20-35 °C), average relative humidity 60% (20-100%) and solar radiation 250 to 300 g.cal.cm-2.día-1.
Five samplings were made every seven days of the aerial and underground
sections of the plants. After 21 days of growth a greater number of leaves and
reduced foliar area were observed as light intensity increased. After 28 and 35
days, the number of leaves, foliar area and length of root decreased, but the
weight of corms increased. Blooming took place at the fifth week. After 65 days
in the 100% light treatment, the number of leaves was higher, while the foliar
area was smaller. The dry weight of inflorescence was smaller at the 25% light
treatment. It is concluded that light is important in the growth of purple nutsedge, since a higher light intensity promoted the
flowering and the number of leaves and larger dry weights of roots, corms and
total. To control purple nutsedge, the application of
an integral method of shade in combination with an herbicide and biological
control is recommended.
Key words: Corms, purple nutsedge, light intensity, leaves
INTRODUCCIÓN
Cyperus rotundus (L.), es
una especie nativa de los trópicos, y con el correr del tiempo se fue
extendiendo hacia áreas subtropicales, de tal forma que actualmente se encuentra en más países, regiones y localidades
que cualquier otra maleza en el mundo (Holm et al 1977).
El corocillo logra su crecimiento en un nivel del mar
hasta cerca de los
Los cormos de C.
rotundus (L.) son blancos, suculentos y casi
redondeados al comienzo de su formación ellos llegan a ser casi negro y muy duros a medida que madura y se
acumulan almidón En pleno desarrollo,
tienen de
El corocillo coquito C. rotundus (L.) es considerado una de las malezas invasoras mas importante del mundo y especialmente de las zonas
tropicales, afectando prácticamente a todas las plantas cultivada (Holm et al, 1977;
Horowitz , 1972). Ninguna otra especie presenta
tantas dificultadas a la producción agrícola, en regiones tropicales y
subtropicales C. rotundus (L.) y todavía a pesar de todo el esfuerzo de
investigación sigue siendo una de las especies más “agresivas” en los sistema
de cultivos y es capaz de interferir dramáticamente e incluso con cultivos como
la caña de azúcar, especies esta que puede alcanzar cuatro metro de altura y
rendir más toneladas que cualquier otro cultivo en el mundo. Puede también
reducir el crecimiento de algunos árboles frutales como es el caso de los
cítricos. Las hortalizas frecuentemente sufren deterioro en calidad, como también pérdidas en rendimiento cuando
los rizomas de C. rotundus
( L.), penetran las raíces del
cultivo, tubérculos o bulbos (Doll, 1983). El C. rotundus (L.),
es considerada una de las plantas dañinas más diseminada agresivamente en todo el
mundo provocando reducciones cualitativamente y cuantitativamente en la
producción mundial en principales cultivos. (Cudney,
1997). Una vez que el corocillo halla infestado un
área es fundamental controlar su diseminación o conseguirlo por medio de cuidado
especiales como son revolviendo el suelo (arado) o revolviendo el suelo y
poderse separar los tubérculos, rizomas, reduciendo la dormancia
y favoreciendo la brotación. Así mismo es importante
adoptar métodos de manejos en los cuales se obtenga un número de disturbios en
el suelo como un sistema de plantación directo en las que ocurre un pequeño
revolvimiento del suelo en las plantas. (Ferreira, 2000).
La luz promueve o inhibe la germinación de semillas de
algunas especies. Las semillas que son inhibidas por la luz son denominadas fotoblásticas negativas (Salisbury y Ross, 1992). La luz es
probablemente el factor ambiental más complejo y variable que actúa sobre las
plantas, desempeñando un papel crucial al proporcionar energía para la
fotosíntesis y actuar como estimulo para el
crecimiento y desarrollo. La fotomorfogénesis
(crecimiento y desarrollo vegetal dependiente de la luz) abarca el conjunto de
procesos mediante los cuales las plantas, adquieren información de la calidad,
cantidad, dirección y fotoperiodicidad de la luz
ambiental que controla su crecimiento y diferenciación (Bergareche y Moysse, 1993). En
las hojas la luz tiene un efecto importante sobre las estomas que es
independiente de la fotosíntesis. Es factible que la luz actúe sobre las células
del mesófilo, las cuales envían algún mensaje a las
células oclusivas, o puede ser que el fotorreceptor
se encuentre en las células oclusivas mismas (Salisbury y Ross, 1992).
No todas las hojas de una planta reciben la misma cantidad de luz, ya que se
hacen sombra unas a otras. Experimentos de laboratorio han demostrado que,
siendo favorables las demás condiciones, la intensidad de la fotosíntesis
aumenta con la intensidad de la luz. En la germinación de las semillas, la
salida del estado de latencia requiere en determinados casos, algunos estímulos
ambientales después de la maduración tales como luz o bajas temperaturas
(García y Primo, 1989). En el campo el manejo de la luz es una de las
herramientas de mayor importancia del Cyperus rotundus (L.), sistema de cultivos, técnicas culturales
pueden ser adoptada para disminuir la cantidad de luz disponibles en plantas
dañinas, pues la calidad e intensidad de luz bajo de las hojas varían con la
naturaleza del cultivo y arreglo espacial de plantas (Mclachan
et al, 1993); por presentar tasa
fotosintética C4, el corocillo es
aptamente eficiente en la asimilación del CO2 atmosférico y en su
conversión a carbohidratos, todavía para que la especie C4 realice con eficiencia la fotosíntesis
ellas necesitan desarrollarse en condiciones de alta temperatura y luminosidad,
por ello el Cyperus
es poca competitiva en
condiciones de bajas temperaturas y/o intensidad
lumínica (Silva, 2000).
Numerosas investigaciones han sido efectuadas sobre el efecto de la luz en el
crecimiento de las plantas, sin embargo, se han encontrado pocos reportes sobre
el efecto de la intensidad de la luz en el crecimiento del C. rotundus (L.). Mediante esta investigación se pudo
determinar algunos efectos de la intensidad de la luz sobre el crecimiento del C. rotundus (L.) bajo tres tratamientos (100, 50 y 25 % de luz).
MATERIALES Y
MÈTODOS
Ubicación:
El presente trabajo se realizó en el
Laboratorio de Fisiología Vegetal del Departamento de Botánica Agrícola de la
Facultad de Agronomía de la Universidad Central
de Venezuela en Maracay estado Aragua, Venezuela-
Material
vegetal
Cormos y plantas de corocillo
Determinación
del porcentaje de
germinación y brotación,
velocidad de germinación y emergencia
Se seleccionaron
aproximadamente 1000 cormos pesándose, por su tamaño y peso similar para
lograr uniformidad de los mismos según
metodología usada por Cárdenas (1992) se
buscaron pesos variados de 1-
Diseño experimental y tratamientos
El diseño estadístico utilizado para determinar el
porcentaje de germinación, brotación, velocidad de
germinación y emergencia de los cormos de Cyperus rotundus L fue completamente aleatorizado azar. Para las
plantas en el invernadero, el diseño usado fue completamente aleatorizado con
tres repeticiones y tres tratamientos de intensidad de luz, 12 plantas por
tratamiento, total 36 plantas. Las intensidades de luz evaluados fueron: 100%
(plena exposición dentro del umbráculo), 50% (una capa de malla de invernadero
blanca) y 25% (dos capas de malla de invernadero blanca). En
Establecimiento de plantas
Una vez germinados los cormos (tres días después de la
siembra) y las plántulas emergidas alcanzaron tres hojas, se sembraron dentro
de recipientes plásticos negros con capacidad para
Determinación de variables
Masa seca total y por órgano (raíces, cormos,
estolones, hojas, flores), área foliar total/planta, área foliar específica,
relación sistema aéreo/sistema radical y desarrollo vegetativo y productivo.
Número de hojas y número de cormos/planta. Inicio de floración, número de
hojas, longitud y ancho de hojas. Las variables
relacionadas con los rendimientos se tomaron una vez efectuadas las
muestras cada siete días durante 35 días.
El material vegetal de las tres plantas por tratamiento, por separado en
hojas, cormos, rizomas, raíces, inflorescencia fue colocado en bolsas de papel
y secado a estufa a 70 ºC por 72 h. para la
determinación de la masa seca.
Para el estudio de las características del sistema
radical se tomó una planta por tratamiento.
Se realizó el muestreo en la fase vegetativa tardía. Se separó el suelo del sistema radical
sumergiendo las bolsas en un recipiente con agua aproximadamente unas
Para medir la longitud radical total se realizó el
método de Newman (1966) modificado por Tennant (1975) el cual consiste en contar el número de
intersecciones entre las raíces y un patrón de líneas que se toman como referencia. Cada sistema radical una vez colocado en
agua fue trasladado a un recipiente de vidrio poco profundo, conteniendo agua
jabonosa y en el fondo se había colocado una cuadrícula de
![]()
El volumen radical se determinó utilizando el principio de desplazamiento
de volúmenes. Se introdujo la raíz en el cilindro graduado lleno de agua y se
midió el volumen desplazado por la muestra de raíces.
El área foliar se determinó multiplicando el largo de la hoja por el ancho
de la hoja
Análisis estadístico
Se uso el procedimiento de Kruskal-Wallis para pruebas no paramétricas en cada una de
las variables de cinco muestreos ejecutados cada siete días. Para evaluar la
normalidad de los parámetros se uso la prueba de Wilk-Shapiro, complementado con el método gráfico del
programa Statistix versión 4.0. Se determinaron las
correlaciones de Pearson y la regresión lineal de las variables muestreadas.
RESULTADOS
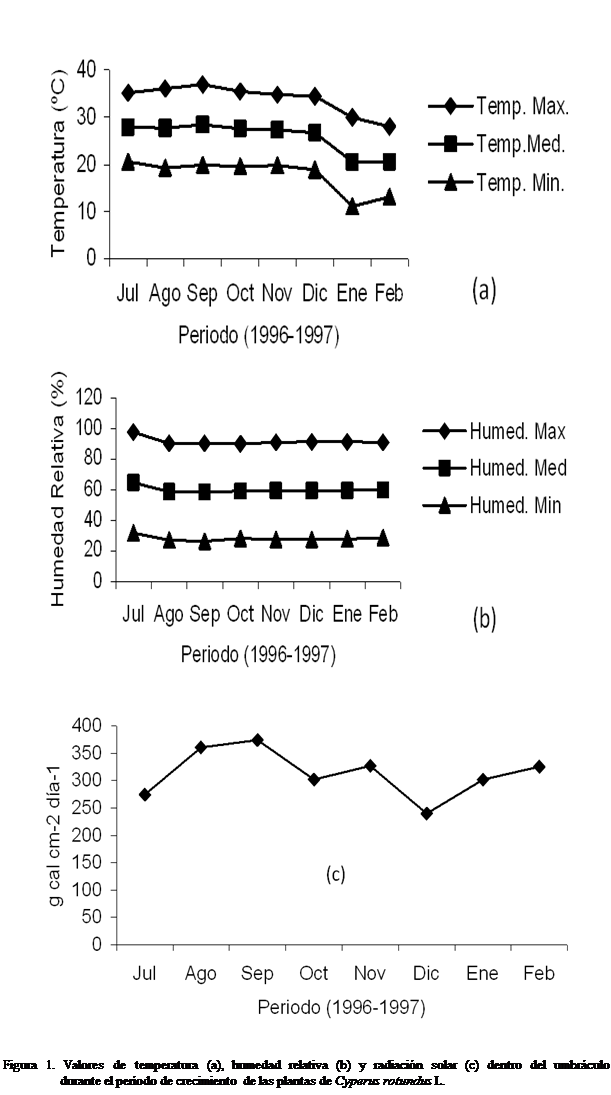
En la figura 1 se pueden observar los valores de temperatura, humedad
relativa y radiación solar dentro y fuera del invernadero durante el tiempo de
realización del experimento. Con los valores registrados se pudo
determinar que durante el crecimiento del corocillo
la temperatura máxima alcanzada fue, por encima de
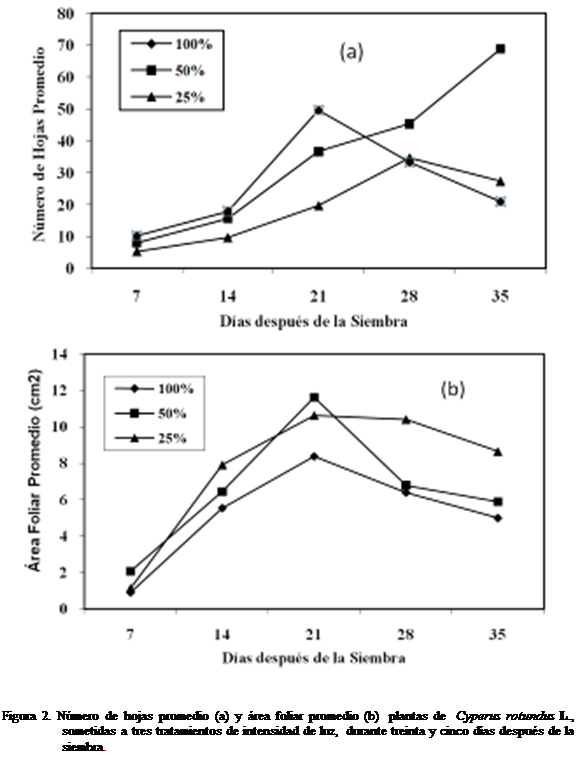
Número de hojas y área foliar
En la figura 2a se observó el número de
hojas mayor a 100% luz a los siete,
catorce y veintiún días después de la siembra., mientras que a los veintiocho y
treinta cinco días fue mayor a 50% luz. El número de hojas a los veintiocho y
treinta y cinco días fue mayor con el tratamiento 25% luz que al alcanzado
con el tratamiento 100% luz. No se observaron
diferencias estadísticamente significativas. En la figura 2b el área foliar a
los siete, y veintiún días después de la siembra, resultó ser mayor con el
tratamiento 50% luz. A los catorce, veintiocho y treinta y cinco días se
observó mayor a 25% luz, siendo menor desde los siete hasta los treinta y cinco
días después de sembrado el corocillo, con el
tratamiento 100% luz. La variable no mostró diferencias estadísticamente
significativas según Kruskal-Wallis.
Longitud de la raíz y volumen de la raíz
En la figura 3a se muestran
los resultados de la longitud de la raíz a los siete días después de la siembra
del corocillo, observándose mayor longitud con
el tratamiento 50% luz que a 25% y
100%. No se observaron diferencias
estadísticamente significativas según Kruskal-Wallis.
Con el tratamiento 50 y 25% luz se observó que
hubo un comportamiento similar en el crecimiento de la longitud radicular a los
catorce días, pero menor al compáraseles con el
tratamiento 100%. A los veintiún días
hubo menor longitud con 25% que a 50%, siendo mayor a un 100% luz.
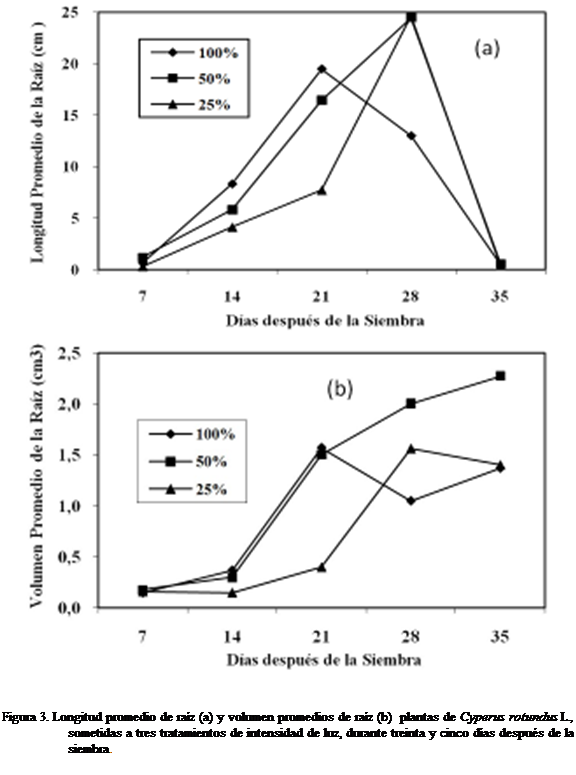
Sin embargo, a los veintiocho
días donde la luz fue menor 25% se obtuvo un pequeño incremento de la longitud
radicular. La longitud radicular obtuvo un crecimiento mayor donde la
intensidad de la luz fue de un 50% a los treinta y cinco días. Al aplicar el
tratamiento 50% luz, el volumen de la raíz se incrementó a partir de los siete
días después de la siembra, hasta los treinta y cinco días (Figura 3b).
Solamente se observó diferencias estadísticamente significativas a los veintiún
días después de sembrado el corocillo según Kruskal – Wallis.
Masa seca (g) de las hojas (PSH) y masa
seca (g) de cormo (PSC)
La masa seca de las hojas (PSH) del corocillo se observó mayor con el tratamiento 50% luz desde
los siete días hasta treinta y cinco después de
la siembra, seguido por el tratamiento 100%, siendo menor a 25% luz (Figura
4a). La masa seca del cormo a los siete días
después de la siembra del corocillo tuvo similar
comportamiento para los tratamientos 100% y 50% luz, siendo menor la masa
alcanzada con el tratamiento 25% luz, a los catorce y veintiún días la masa seca del cormo fue
mayor a un 50% luz (Figura 4b). A los veintiocho días de haber sido sembrado el
Cyperus rotundus
L., la masa seca del cormo fue mayor
a un tratamiento 25% luz que a 50% y
100% luz, alcanzando ser menor con el tratamiento 100% luz. A los treinta y cinco días de siembra la masa
seca del cormo del corocillo, tuvo variación
observándose a éste mayor con un 50% luz, seguido del tratamiento 100% luz y
menor a un 25% luz (Figura 4b). Las dos variables masa seca de la hoja (PSH) y masa seca del cormo (PSC) no mostraron
diferencias estadísticamente significativa según Kruskal-Wallis.
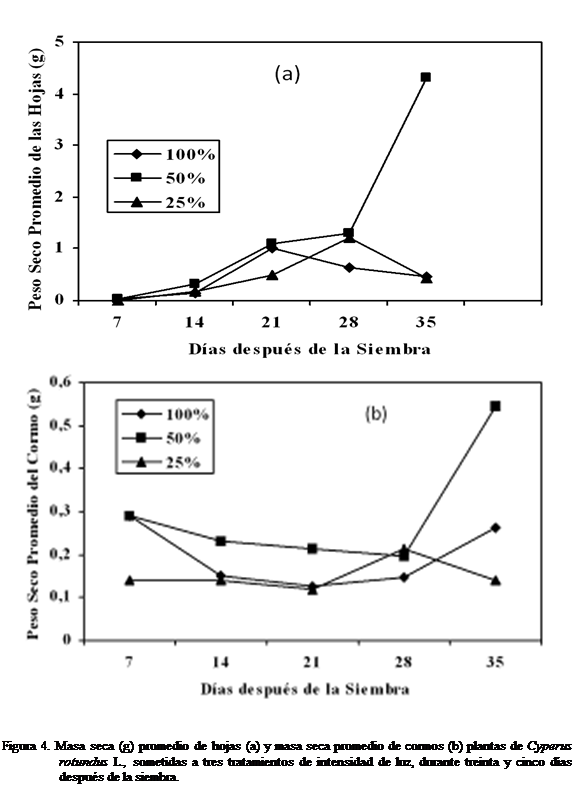
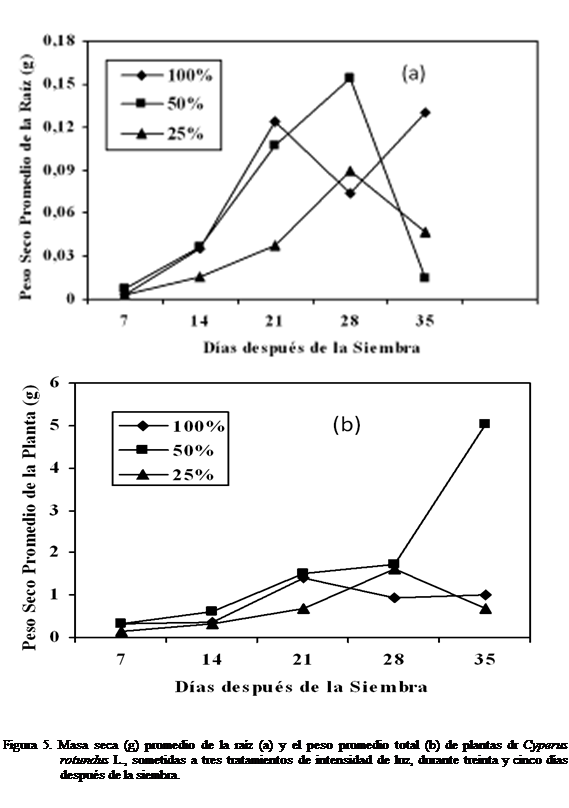
Masa seca (g) de la raíz (PSRA) y masa seca (g)
total (PST)
La
masa seca de la raíz (PSRA) del corocillo a los siete y catorce días después de la siembra se observó mayor bajo tratamiento
50% luz (Figura 5a). Hubo diferencias estadísticamente significativas según Kruskal-Wallis. A los veintiún días fue mayor con el
tratamiento 100% luz, observándose también diferencias significativas. A los
veintiocho días después de sembrado el Cyperus
rotundus L, la masa seca de la raíz continuó
siendo mayor con el tratamiento 50% pero menor con el 100%. No se observaron
diferencias estadísticamente significativas, mientras que a los treinta y cinco fue menor con el
tratamiento 50%, y mayor con el 100% luz (Figura 5a). La mayor masa seca total a los siete, catorce,
veintiún, veintiocho y treinta y cinco días después de la siembra del Cyperus rotundus
L, se alcanzó con el tratamiento 50% luz, y el menor con el
tratamiento 25%, no observándose diferencias estadísticamente significativas
(Figura 5b).
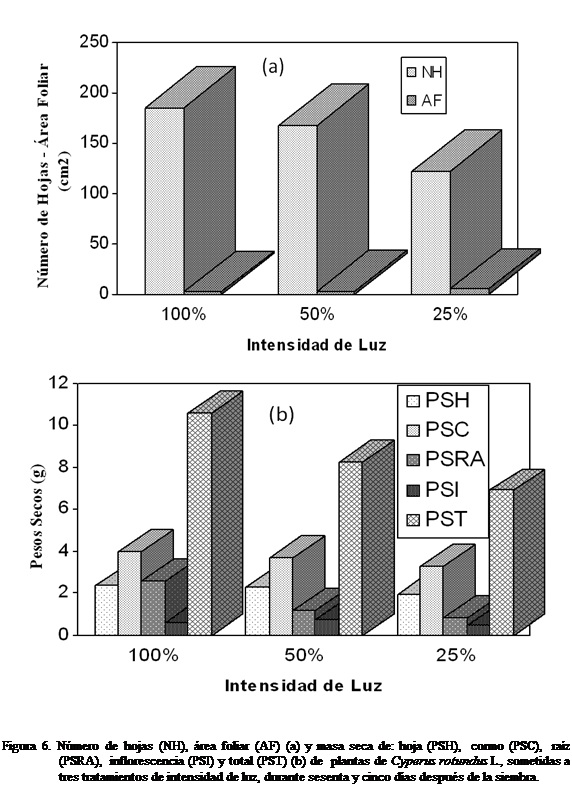
Número de hojas (NH), longitud de hojas (LH),
anchura de hojas (AH), área foliar (AF), masa seca de la hoja (PSH), masa seca
del cormo (PSC), masa seca de la raíz (PSRA), masa seca inflorescencia (PSI) y
masa seca total (PST)
A los
sesenta y cinco días después de la siembra
se puede observar el mayor número de hojas con el tratamiento 100% luz y menor con 25%,
obteniéndose diferencias estadísticamente significativas según Kruskal-Wallis
(Figura 6a). El área foliar fue mayor con el tratamiento 25% luz, no
observándose deferencias significativas (Figura 6a). La masa seca de la hoja,
cormo, raíz y total se observaron mayor a una intensidad de luz 100%, mientras que la masa seca de la
inflorescencia fue menor, siendo mayor bajo tratamiento 50% luz (Figura 6b). No
hubo diferencias estadísticamente
significativas.
DISCUSIÓN
El C. rotundus (L.) es la
maleza más dañina del mundo (Holm et al, 1977). Ha sido bien clasificada por sus características
morfológicas. Los resultados obtenidos con los cormos germinados sobre papel
toalla en bandejas plásticas, dentro de las gavetas de los escritorios no
fueron afectados aparentemente para su brotación
coincidiendo con lo señalado por ( Cárdenas, 1992) cuando menciona que los
tubérculos aislados en cualquier posición independientemente de su tamaño
pueden germinar y producir brotes. El porcentaje de germinación de los cormos
en bandejas encima de los escritorios fue menor, posiblemente debido a la luz y humedad (Cárdenas, 1992) señala que el crecimiento de
las yemas en tubérculos de corocillo es afectado
fuertemente por la luz y la humedad.
Durante el crecimiento del corocillo bajo
tratamientos 100, 50, y 25 % luz, en el invernadero, la temperatura máxima
alcanzada estuvo, por encima de
Los resultados observados durante los tres
primeros muestreos efectuados a los siete, catorce y veintiún días después de
la siembra nos demuestran que a mayor intensidad de luz mayor es el número de
hojas y área foliar. A mayor número de
hojas menores fueron las masas secas totales
alcanzadas. A los veintiocho y treinta y cinco días después de la siembra, a
mayor intensidad de luz hubo una disminución en el número de hojas, por lo
tanto las masas secas de las partes subterráneas y aéreas del C. rotundus
(L.), resultaron menores. (Hauser, 1962) menciona que
la masa seca de la raíz, rizoma y masa seca de los cormos, generalmente
aumentan con un incremento en la intensidad de luz y el tiempo. (Hauser, 1962), la floración comienza al mismo tiempo
que se comienzan a formar los tubérculos
después de seis semanas, Doll, (1983) la floración
ocurre a partir de las
CONCLUSIONES
1.
Los cormos
colocados sobre toallas humedecidas y etiolados
germinaron en un 100%.
2.
Los cormos
colocados en toallas humedecidas y sujetos a condiciones de luz, humedad y
temperatura ambiente dentro del laboratorio, germinaron aproximadamente en un
70%, posiblemente debido a la luz y a una menor humedad.
3.
La masa seca de
la inflorescencia fue menor a los sesenta y cinco días en el tratamiento con
menor intensidad de luz (tratamiento 25 %).
4.
A mayor
intensidad de luz (tratamiento 100%) se estimuló en las plantas de corocillo la floración a los 35 días después de la siembra.
5.
Una vez emitida la
floración en el tratamiento 100% luz a los 35 días después de la siembra,
ocurrió una disminución de la longitud de la raíz y del área foliar e
incrementó la masa seca de los cormos.
6.
A mayor número de
hojas menores fueron las masas secas totales
alcanzadas.
7.
A mayor
intensidad de luz (tratamiento 100%) mayores fueron el número de hojas y la
masa seca de las hojas, raíces, cormos y total a los 65 días después de haberse
sembrado el corocillo.
LITERATURA CITADA
Arnaude, O. 1994.
Influencia del stress por fósforo sobre el desarrollo y la respiración radical
en caraota (Phaseolus vulgaris)
cv. Manaure, frijol (Vigma ungiculata) cv. Tuy, tomate (Lycopersicon
sculentum Mill), Euphorbia heterophyla L.
y Amaranthus
dubius Mart. Trabajo
de grado para Magister Scientiarium en Agronomía UCV. Facultad de Agronomía.
Inst. Bot. Agri.
Maracay.179 pp.
Bell, R. S.; W. H. La Chman, E. M. Rhan and R. D. Sweet. 1962.
Life history studies as related to weed control in the northeast. I. Nutgrass. Rhode
Island Agric. Exp. Stn. Bull N°364.
33 p.
Bergareche, C. y L. Moysse. 1993. Illuminating Phytochrome
Functions there is light al the end of the
Tunnel. Plant Physiol. 393-394.
Cárdenas, C. H. 1992. El corocillo (Cyperus rotundus L
):
Cudney, D. W. 1997. Nutsedge:
hystory, economy, importance and distribution. In:
NUTSEDGE Management Workshop. Reverside, University
of California, 1997. (http://www.cnas.ucr.edu/-bps/hnutsedge.htm.)
Doll, J. 1983. Yellow Nutsedge
Control in Field Crops. University of
Wisconsin Extension Bulletin Nº A2990. 4 pp.
Ferreira, F.A. 2000. Manejo
integrado de plantas daninhas
García, A. P. and M. E. Primo. 1989. Ultraestructural
and biochemical changes in cotyledon
reserve tissues during
germination of Citrus seeds. Journal Experimental Botany. 40:
387-390.
Hauser, E. 1962. Development of purple nutsedge under field conditions. Weeds 10: 315-321.
Holm, L. G.; D. L. Plucknett, J.
V. Pancho and J. P. Herberger. 1977. The World’s Worst
Weeds. University of Hawaii Press. 609 p.
Jha, P.
and D. Sen. 1980. Drought Avoidance by Cyperus
rotundus. Folia Geobotanica Phytotaxonomica 15:
387-394.
Kolesnikov, V. A. 1975.
The root systems of fruit plant. Mir. Publishers, Moscú.
Traducción Ludmila Akesno. 268 p.
Keeley, P. and R. Thullen.
1978. Light requirements of
yellow nutsedge (Cyperus esculentus)
and Light Interceptions by Crops .Weed Sci. 26: 10-16.
McLachlan, S. M.; M. Tollenaar, C. J. Swanton
and S. F. Weise 1993. Effect of corn induced shading on dry matter
accumulation, distribution, and architecture of redroot pigweed (Amaranthus retroflexus).
Weed Sci. 41: 568-573.
Newman, E.L.
Salisbury, I. B. and C .W.
Ross. 1992. Plant Physiology. Wadsworth. Publishing, California, USA.682 p.
Silva, A. A. 2000. Controle de plantas daninhas. In: Associação Brasileira de Educação Agrícola Superior. Curso de proteção de plantas. Brasília, D.F., 2000. p. 133-172.
Tennant, D. 1975. A test of a modified line
intersect method of estimating root length. J. Ecol.(63) 995- 1001.
William, R. 1978. Photoperiod effects on the reproductive
Biology of Purple nutsedge (Cyperus rotundus).
Weed Sci. 26: 539-592.
Wills, G. and G. Briscone. 1970. Anatomy of
purple nutsedge. Weed Sci. 18: 631-635.
Página diseñada por Prof. Jesús Rafael Méndez Natera
TABLA DE CONTENIDO DE LA REVISTA CIENTÍFICA UDO
AGRÍCOLA


