
Revista Científica UDO Agrícola Volumen 8.
Número 1. Año 2008. Páginas: 61-66
Relación entre la tasa de imbibición y el
porcentaje de germinación en semillas de maíz (Zea mays L.), caraota (Phaseoulus
vulgaris L.) y quinchoncho (Cajanum cajan (L.) Mill.)
Relationship
between imbibition rate and seed germination of corn, French bean and pigeonpea
seeds
Jesús
Rafael MÉNDEZ NATERA![]() 1, José Fernando MERAZO PINTO1 y
Nelson José MONTAÑO MATA1
1, José Fernando MERAZO PINTO1 y
Nelson José MONTAÑO MATA1
1Departamento de Agronomía, Escuela de Ingeniería
Agronómica, Núcleo Monagas, Universidad de Oriente, Campus Los Guaritos, Avenida Universidad, Maturín, 6201, edo. Monagas, Venezuela.
E-mail:
jmendezn@cantv.net ![]() Autor para
correspondencia
Autor para
correspondencia
|
Recibido: 25/06/2008 |
Fin
de primer arbitraje: 22/08/2008 |
Primera
revisión recibida: 30/08/2008 |
|
Fin
de segundo arbitraje: 18/09/2008 |
Segunda
revisión recibida: 05/10/2008 |
Aceptado: 21/10/2008 |
RESUMEN
El objetivo
fue determinar la relación entre la tasa de imbibición y el porcentaje de
germinación de semillas de maíz (Zea mays
L.) tratadas y no tratadas con vitavax (carboxin), caraota (Phaseoulus vulgaris L.) y quinchoncho (Cajanum cajan (L.) Mill.). Se pesaron
dos lotes con 10 semillas cada uno y se sometieron a diferentes periodos de
imbibición (1, 4, 8, 12 y 14 horas). La tasa de imbibición se calculó restando
el peso de las semillas después de la imbibición del peso inicial. Para el
porcentaje de germinación de las semillas imbibidas se tomaron 100 semillas de
los cultivos anteriores y se colocaron en tres toallas de papel absorbente y
luego se cubrieron con otra toalla y posteriormente se humedecieron. Luego se
enrollaron las toallas y se colocaron en el germinador a 25 °C. La tasa de imbibición de las semillas de maíz
tratado, caraota y quinchoncho tuvieron un comportamiento cuadrático y la mayor
tasa ocurrió a los 8, 10 y 10 horas, respectivamente. La respuesta de las
semillas de maíz sin tratar fue irregular. La germinación de las semillas de
maíz sin tratar disminuyó con incrementos de los tiempos de imbibición, similar
tendencia tuvieron las semillas de quinchoncho, mientras que para las semillas
de caraota disminuyó hasta las 8 horas para luego incrementarse, la germinación
de las semillas de maíz tatado fue muy baja (< 12%). No se encontró una
relación entre la tasa imbibición y los porcentajes de germinación en ninguno
de los cultivos estudiados.
Palabras clave:
Germinación, imbibición, Zea mays, Phaseoulus vulgaris, Cajanum cajan
ABSTRACT
The
objective was to determine the relationship between imbibition rata and
germination percentage of seed of maize (Zea
mays L.) treated and untreated with vitavax (Carboxin), French beans (Phaseoulus vulgaris L.) and pigeonpea (Cajanum cajan (L.) Mill). Two lots were
weigthed with 10 seeds each one and they were subjected to different imbibition
periods (1, 4, 8, 12 and 14 hours). Imbibition rate was calculated as: seed
weight of imbibited seeds minus initial seed weight. One hundred seeds were
taken from previous crops and placed on three towel papers and covered with one
towel paper and then they were moistened to determine germination percentage of
different imbibited seeds at different periods. The towel papers were rolled
and placed on the germination cabinet at 25 ºC. Imbibition rate of treated
corn, French bean and pigeon pea presented a quadratic behavior and the bigest
rate ocurred at 8, 10 and 10 hours, respectively. The performance of untreated
corn seeds was irregular. The seed germination of treated corn decreased as
increased imbibition period, similar trend was presented by pigeon pea seed
germination, while French bean germination decresed from 1 to 8 hours of
imbibition, after that, the germination was increased. The seed germination of
treated corn was very low (< 12%). There was not a relationship between
imbibition rate and seed germination in the crops.
Key words:
Germination, imbibition, Zea mays, Phaseoulus vulgaris, Cajanum cajan
INTRODUCCIÓN
La semilla es esencial para la supervivencia de la
humanidad, por cuanto almacena el más alto potencial genético que la ciencia
pudiera llegar a desarrollar y es un elemento vital en la agricultura moderna,
la semilla certificada contribuye a alcanzar una producción más alta (Douglas
1991).
El agua es esencial para la
rehidratación de las semillas, siendo el paso inicial antes de la germinación.
La cantidad de agua absorbida por una semilla imbibida depende de un número de
factores, por ejemplo, el tamaño, hidratabilidad de los contenidos, etc., pero
en términos absolutos, la absorción de agua es relativamente pequeña y a menudo
puede no exceder
La geminación se inicia con la imbibición y termina
con la emergencia. La imbibición es la toma de agua por parte de la semilla
seca, sin importar si ésta se encuentra viable o no, y la emergencia es el
proceso por el cual el eje embrionario en especies dicotiledóneas o radícula en
monocotiledóneas crece, se extiende y atraviesa las estructuras que lo rodean
(Azcón y Talon, 2003). La absorción de agua por parte de la semilla está
directamente influenciada por la presencia de la testa y la permeabilidad que
ésta tenga. El tejido de reserva absorbe agua a una velocidad intermedia hasta
completar su hidratación (Moreno et al.,
2006).
La germinación de las semillas puede
ser definida como una serie de procesos metabólicos y morfogenéticos, los
cuales resultan en la transformación del embrión en una plántula capaz de
convertirse en una planta adulta. Durante la germinación ocurre procesos
secuenciales y sincronizados y son reconocidos de tal manera que los procesos
anabólicos y catabólicos toman lugar de manera simultánea. (Coll et al., 1995). Muchas semillas colocadas
en agua destilada en cápsulas de Petri bajo condiciones óptimas para la
germinación muestran un patrón trifásico de absorción de agua. La absorción
inicial de agua en
Powell y Matthews (1979) obervaron
consistentemente que la rápida absorción de agua por los embriones secos en
arveja efectivamente matan las áreas de los tejidos sobre las superficies
abaxiales de los cotiledones. Retardando la absorción de agua bien sea dejando
la testa o imbibiendo en una solución al 30 % de polietilenglicol 4000
(Carbowax) previene la producción de área muertas. Este fenómeno, en la cual
las áreas de células muertas son producidas por la rápida absorción de agua, se
conoce ahora como daño de la imbibición. Aunque las semillas imbibidas en agua
a temperaturas bajas (
La testa claramente es una barrera
para la absorción de agua en las leguminosas la cual sirve para proteger al
embrión seco del efecto de daño causado por una rápida absorción de agua, un
rol de un considerable valor adaptativo. Esta concepción da una mayor
significancia a las rupturas en la testa, porque ellas pueden facilitar la
absorción de agua y permitir que ocurra una rápida imbibición, con los
consecuentes efectos sobre los cotiledones y el rendimiento de la semilla.
Varias características de embriones que han sufrido daño por imbibición fueron
la reminiscencia de semillas de bajo vigor. La habilidad de germinación
permaneció alta, las áreas de tejido muerto se encontraron en las superficies
abaxiales de los cotiledones imbibidos, la retención de soluto fue pobre y los
embriones exhibieron una respiración reducida y un lento desarrollo de las
plántulas (Matthews et. al. 1980).
Oguntunde y Adebawo (1989)
determinaron los patrones individuales de absorción de agua de semillas de maíz
cv ‘TZPB’, maíz blanco local, tipos blancos y rojos de sorgo y millo a 30,
35, 40 y 45 °C remojadas durante 0 a 72 horas. Las curvas
de hidratación obtenidas mostraron que la mayor absorción de humedad de las
semillas de estos materiales ocurrió dentro de las primeras 24 horas, mientras
que la máxima absorción de agua o contenido de saturación de humedad ocurrió
aproximadamente a las 36 horas, sin importar la temperatura en el remojo. El
contenido de humedad máximo se incrementó cuando la temperatura en el remojo
subió de 30 a 45 ºC.
En estudios realizados para mejorar
la calidad de la semilla con el uso de tratamientos tales como condicionamiento
osmótico, humedecimiento inicial y el uso de biorreguladores, es importante
obtener información de las etapas de absorción de agua para diferentes especies
(Zambello de Pinho et al. 2004). El
objetivo fue determinar la relación entre la tasa de imbibición y el porcentaje
de germinación en semillas de maíz, caraota y quinchoncho.
MATERIALES Y MÉTODOS
Se pesaron dos lotes con 10 semillas
cada uno y se sometieron a diferentes periodos de imbibición (1, 4, 8, 12 y 14
horas). La tasa de imbibición se calculó restando el peso de las semillas
después de la imbibición menos el peso inicial. Para el porcentaje de
germinación de las semillas imbibidas se tomaron 100 semillas de los cultivos
anteriores y se colocaron en tres toallas de papel absorbente y luego se
cubrieron con otra toalla y posteriormente se humedecieron. Luego se enrollaron
las toallas y se colocaron en el germinador a 25 °C. Se realizó el análisis de regresión de la tasa de
imbibición y el porcentaje de germinación en función de los tiempos de
imbibición (Draper y Smith, 1998). Finalmente se determinó el coeficiente de correlación
de Pearson entre estos dos caracteres (Steel et al. 1996).
RESULTADOS
En el cuadro 1 se muestran los
resultados para la tasa de imbibición y el porcentaje de germinación.
|
Cuadro
1. Tasa de imbibición (TI) y porcentaje de germinación (% G) de semillas de
maíz (Zea mays L.), caraota (Phaseoulus vulgaris L.) y quinchoncho (Cajanum cajan (L.) Mill.), |
||||||||
|
Horas de |
Caraota |
Maíz Tratado |
Maíz Sin Tratar |
Quinchoncho |
||||
|
Inmersión |
TI |
% G |
TI |
% G |
TI |
% G |
TI |
% G |
|
14,00 |
0,60 |
100,00 |
0,00 |
7,00 |
0,30 |
93,00 |
0,25 |
79,00 |
|
12,00 |
1,35 |
67,00 |
0,45 |
3,00 |
0,09 |
84,00 |
0,37 |
67,00 |
|
8,00 |
1,35 |
65,00 |
0,51 |
0,00 |
0,05 |
95,00 |
0,63 |
88,00 |
|
4,00 |
0,48 |
60,00 |
0,48 |
13,00 |
0,21 |
100,00 |
0,15 |
85,00 |
|
1,00 |
0,00 |
87,00 |
0,00 |
1,00 |
0,15 |
100,00 |
0,20 |
83,00 |
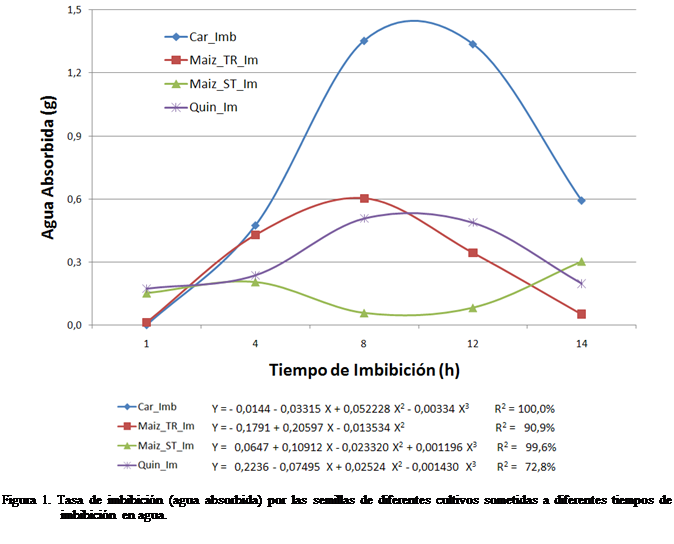
La figura 1 muestra la relación entre la tasa de
imbibición y el tiempo de imbibición. Se observa que las máximas tasas de
absorción de agua ocurrieron a las 8, 10 y 10 horas después de la imbibición
para los cultivos maíz tratado, caraota y quinchoncho, respectivamente, en el
caso del maíz sin tratar la mayor tasa
ocurrió a las 14 horas, aunque la respuesta fue irregular.
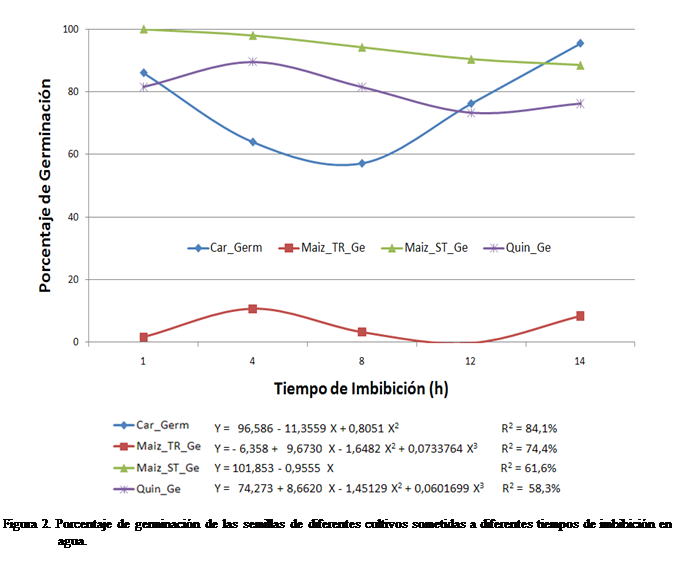
La Figura 2 muestra
la relación entre el porcentaje de germinación y los periodos de imbibición.
Para el maíz sin tratar se observó que a medida que se incrementó el periodo de
imbibición, la germinación fue menor, similar tendencia tuvieron las semillas
de quinchoncho aunque la germinación fue máxima a las 4 horas de imbibición.
Por otra parte, la germinación de las semillas de caraota fue menor a las 4 y 8
horas de imbibición para luego incrementar en los dos mayores periodos de
imbibición. La germinación del maíz tratado fue muy baja y siguió una tendencia
irregular.
En el cuadro 2 se observa el coeficiente de
correlación entre la tasa de imbibición y el porcentaje de germinación dentro de cada
cultivo. No se observó ninguna asociación significativa entre estos dos
caracteres, aunque el coeficiente de correlación fue de -0,49 para el cultivo
de caraota, sugiriendo un posible efecto negativo de la tasa de absorción de
agua sobre la germinación de las semillas de caraota.
|
Cuadro 2. Coeficientes de correlación de Pearson entre la tasa de imbibición
y el porcentaje de germinación dentro de cada cultivo. |
||||
|
Car_Ger |
MT_Ger |
MsT_Ge |
Qui_Ge |
|
|
Car_Imb |
-0,49 |
|||
|
MT_Imb |
0,12 |
|||
|
MsT_Imb |
0,26 |
|||
|
Qui_Imb |
0,11 |
|||
|
Car_, MT_,
MsT_ y Qui: Semillas de Caraota (Phaseoulus
vulgaris L,), maíz (Zea mays
L.) tratado, maíz sin tratar y quinchoncho
de maíz (Zea mays L.),
caraota (Phaseoulus vulgaris L.) y
quinchoncho (Cajanum cajan (L.)
Mill.), Imb: Tasa de
imbibición y Ge: Porcentaje de germinación Todos los coeficientes de correlación fueron no
significativos (p > 0,05). |
||||
DISCUSIÓN
Se observó que a medida que las semillas están más
tiempo sumergidas en agua, estas imbiben más agua y por consiguiente aumenta la
tasa de imbibición y luego disminuyó esta tasa, esto se debe a que las semillas
imbiben agua hasta cierto tiempo, cuando disminuye abruptamente la absorción de
agua, para luego reiniciarse la absorción, generando una curva con tres fases
bien definidas, la fase I de la imbibición es cuando las semillas absorben agua
en una forma proporcional al tiempo (Bewley y Black 1983) lo cual fue observado
en este ensayo para los cultivos maíz tratado, caraota y quinchoncho con
incrementos de las tasas hasta las 8, 10 y 10 horas después de la imbibición,
respectivamente. Sousa Paiva et al.
(2006) indicaron que durante la imbibición, el incremento del peso de las
semillas de Swietenia macrophylla fue
considerable, aunque la alteración en el volumen fue discreta. En este experimento
se observó el mismo patrón de la curva de imbibición pero sólo para las dos
primeras fases, es decir, la primera de absorción rápida para luego no absorber
agua por un cierto tiempo, la tercera fase no ocurrió debido a que el ensayo
terminó cuando comenzaba la misma (protrusión de la radícula).
Resultados análogos fueron reportados por Ogutande y
Adebawo (1989) quienes encontraron que la absorción de agua en el cultivar TZPB
(semilla blanca) ocurrió dentro de las primeras 24 horas. Similarmente, Mei y
Song (2008) estudiaron los eventos iniciales tanto morfológicos como
fisiológicos que ocurren durante la imbibición y germinación de semillas de
maíz e indicaron que la absorción de agua de las semillas exhibió un patrón
trifásico con un incremento marcado durante la fase inicial de imbibición y
luego una disminución lenta, seguido por un segundo incremento sustancial. El
tiempo de imbibición para 10 y 50% de la germinación de las semillas fue de
aproximadamente 26 y 46 horas a 30 ºC, respectivamente. Sousa Paiva et al. (2006) indicaron que durante la
imbibición, el movimiento de agua dentro de la semilla se debe a la acción de
difusión y capilaridad, con el movimiento del agua de una región con un
potencial hídrico mayor a una de un potencial hídrico menor.
En general, las semillas de maíz sin
fungicida tuvieron el mayor porcentaje de germinación, seguidas de las semillas
de quinchoncho y caraota, finalmente el maíz con vitavax, este último con
porcentajes de germinación menores al 20 %, en comparación al maíz sin vitavax
que tuvo porcentajes superiores al 80 % en los diferentes tiempos de inmersión.
Almeida Pinto (2000) estudió la eficiencia de los fungicidas captan (120, 90
and 60 g de ingrediente activo/100 kg), thiram (140), thiabendazole (20),
thiram + thiabendazole (75 + 10), and carboxin + thiram (75 + 75) en la
germinación de semillas de maíz del cultivar BR 106 y encontró que ningún
fungicida presentó fitotoxicidad en las semillas. Stankovich y Pavlov (1987a,
1987b) trataron semillas de 5 líneas de maíz con los fungicidas thiram y captan
y encontraron que el porcentaje de germinación fue mayor que el control no
tratado (excepto para Mo17 con captan y B312 con thiram), esto sugiere que los
resultados encontrados en este ensayo pudieron deberse entre otras cosas a que
la dosis aplicada fue demasiado alta. Ramírez (1975) señala que pocas
sustancias causan daño al poder germinativo de la semilla.
CONCLUSIONES
La tasa de imbibición de las semillas de maíz tratado,
caraota y quinchoncho tuvieron un comportamiento cuadrático y la mayor tasa
ocurrió a los 8, 10 y 10 horas, respectivamente. La respuesta de las semillas
de maíz sin tratar fue irregular. La germinación de las semillas de maíz sin
tratar disminuyó con incrementos de los tiempos de imbibición, similar
tendencia tuvieron las semillas de quinchoncho, mientras que para las semillas
de caraota disminuyó hasta las 8 horas para luego incrementarse, la germinación
de las semillas de maíz tatado fue muy baja (< 12%). No se encontró una relación
entre la tasa imbibición y los porcentajes de germinación en ninguno de los
cultivos estudiados.
LITERATURA CITADA
Almeida Pinto, N. F. J de.
2000. Tratamento fungicida de sementes de milho contra fungos do solo e o
controle de Fusarium associado às sementes. Scientia Agricola 57 (3): 483-486.
Azcón Bieto J. y M. Talón.
2003. Fundamentos de Fisiología Vegetal. McGrawHill/Interamericana. Barcelona,
España. 522 p.
Bewley, J. D. and M. Black. 1983. Physiology and Biocemistry of Seeds in
relation to germination. Volume 1: Development, germination, and growth. Second
edition. Berlin, Germany. Springer-Verlag. 306 p.
Coll, J. B.; G. N. Rodrigo, B.
S. Garcia and R. S. Tamés. 1995. Fisiología vegetal. Madrid. Ediciones
Pirámide. 662 p.
Douglas, J. E. (comp., ed.).
1991. Programas de semillas. Guia de planea-ción y manejo. Centro Internacional
de Agricultura Tropical (CIAT). Segunda edición. Cali, Colombia. 358 p.
Draper, N. R. and H. Smith.
1998. Applied regression analysis. Third edition. John Wiley & Sons. New
York, USA. 736 p.
Matthews, S.; A. A. Powell and
N. E. Rogerson. 1980. Physiological aspects of the development and storage of
pea seeds and their significance to seed production. In Seed Production (ed. P.
D. Hebblethwaite). Butterworths, London, England. p. 513-525.
Mei, Y. and S. Song. 2008.
Early morphological and physiological events occurring during germination of
maize seeds. Agricultural Sciences in China 7 (8): 950-957.
Moreno, F.; G. A. Plaza y S.
V. Magnitskiy. 2006. Efecto de la testa sobre la germinación de semillas de
caucho (Hevea brasiliensis Muell.).
Agronomía Colombiana 24 (2): 290-295.
Oguntunde, A. C. and O. O.
Adebawo. 1989. Water-uptake pattern during traditional soaking of cereal
grains. Tropical Science 29 (3): 189-197.
Powell, A. A. and S. Matthews.
1979. The influence of testa condition on the imbibition and vigour of pea
seeds. Journal of Experimental Botany 30: 193-197.
Ramírez G, M. 1975.
Almacenamiento y conservación de granos y semillas. Editorial Continental, S.
A. México. 300 p.
Sousa Paiva, E. A.; J. P.
Lemos-Filho and D. M. Trombert Oliveira. 2006. Imbibition of Swietenia macrophylla (Meliaceae) seeds:
The role of stomata. Annals of Botany 98 (1): 213-217.
Stankovich, M. and M. Pavlov.
1987a. Test of the efffect of the fungicide TMTD and captan on the initial
growth of inbred maize lines. Informatsionnyi Byulleten´poKukuruze. No. 6,
307-323.
Stankovich, M. and M. Pavlov.
1987b. Testing de effect of the fungicide TMTD and captan on early growth of
inbred maize lines. Arhivza Poljoprivredne Nauke 48 (170): 161-172.
Steel, R. G. D.; J. H. Torrie
and D. A. 1996. Principles and procedures of statistics: A biometrical
approach. Third sub edition. McGraw-Hill Company. New York, USA. 672 p.
Zambello de Pinho, S.; L. R.
de Carvalho e M. E. Aparecida Delachiave. 2004. Limit between stages I and II
of a seed imbibition curve. Sci. Agric. 61 (1): 17-20.
Página diseñada por Prof. Jesús Rafael Méndez Natera
TABLA DE CONTENIDO DE LA REVISTA CIENTÍFICA UDO
AGRÍCOLA


