
Revista Científica UDO Agrícola Volumen 7. Número 1. Año 2007. Páginas: 274-284
Mesozooplankton composition and distribution in relation to oceanographic conditions in
the Gulf of Cádiz,
Composición y distribución del mesozoopláncton
en relación a condiciones oceanográficas en el Golfo de Cádiz, España
Paulo MAFALDA Jr.1 ![]() , Juan PÉREZ DE
RUBÍN2 and Christiane SAMPAIO DE SOUZA1,3
, Juan PÉREZ DE
RUBÍN2 and Christiane SAMPAIO DE SOUZA1,3
1Universidade
Federal da Bahia (UFBA). Instituto de Biologia. Laboratório de Plâncton.
40.210-020. Salvador. BA. Brasil. 2Instituto Español
de Oceanografía (IEO). Muelle
pesquero s/n Apartado 285.29640. Fuengirola/MA,
España. 3Universidade Federal da Bahia
(UFBA). Instituto de Geociências. Curso de Pós-Graduação em Geologia.
40.210-020. Salvador. BA. Brasil. E-mails: pomafa@ufba.br and
jprubín@ma.ieo.es ![]() Corresponding author
Corresponding author
|
Received: |
First reviewing ending: |
First review received: |
|
Second reviewing ending: 09/24/2007 |
Second review received: |
Accepted: |
ABSTRACT
Two surveys were conducted during July 1994 and July
Key words: Mesozooplankton, cladocera, biomass, spatial distribution,
RESUMEN
Dos
campañas de verano fueron realizadas en los meses de julio de 1993 y de 1994 en
el Golfo de Cádiz, con el objetivo de discernir
patrones temporales y espaciales en la biomasa y composición del mesozoopláncton y investigar su
relación con las condiciones oceanográficas. Variabilidad temporal entre los
dos veranos ha sido observada. La columna de agua ha sido más caliente y salina
en 1994 y también ha presentado una mayor densidad y biomasa de mesozoopláncton. A pesar de estas variaciones temporales la
presencia de varias especies planctónicas fueron notablemente regulares. En los
dos veranos los cladóceros (Penilia avirostris, Evadne spinifera,
Evadne tergestina,
Evadne nordmanni,
y Podon spp) fueron el grupo más abundante seguido por los copépodos y apendicularias. La dominación de cladóceros en verano es
básicamente debido a la alta abundancia de Penillia avirostis. El estudio de
tendencias demostró que la abundancia relativa de copépodos aumentó tras los
veranos, aunque este aumento solamente ha sido significativo en las estaciones
occidentales, donde es más elevada la influencia del agua Atlántica. La
densidad de mesozoopláncton and Cladocera
demostró una correlación positiva con temperatura y biomasa pero demostró una
correlación negativa con salinidad y profundidad. La biomasa y la densidad de Copepoda y Appendiculata no han presentado correlación con las variables oceanográficas.
Palabras
clave: Mesozooplancton,
cladocera, biomasa, distribución espacial, Golfo de
Cádiz
INTRODUCTION
The

Surprisingly,
the interest attracted by fisheries exploitation in
this area was not followed by research aimed to characterize the mesozooplankton community, a key factor to the consequent
fish larval survival and the fisheries yield.
The mesozooplankton plays an important role in the marine food
web as a link between the micro- and macrozooplankton
(Neumann-Leitão et
al., 1999). However, little is known about the spatial and temporal
variability of zooplankton on the shelf in the
The
area of the Gulf of Cádiz is characterized by an ample continental shelf,
around
This paper presents results of the meso-scale mesozooplankton
composition and its spatial and temporal variability in the
MATERIAL AND METHODS
During
July 1994 and July
Results AND DISCUSSION
Oceanographic
conditions
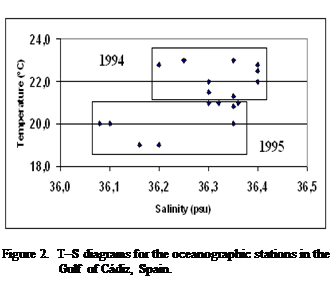
According
to the T-S diagram (Figure 2), there were significant differences in
environmental conditions in the surface water between the two summers (Table
1). The upper water column was warmer (p < 0.05) and more saline (p <
0.05), in 1994. The water column was generally well stratified and the
thermocline was located on average at
|
Table 1. Oceanographic
variables (mean ± SD) during summers in
the |
|||
|
|
1994 |
1995 |
Welch t Test |
|
Depth
(m) |
63.60
± 34.4 |
63.10
± 34.7 |
P = 0.9708 |
|
Salinity
(psu) |
36.33
± 0.06 |
36.27
± 0.11 |
P = 0.0183* |
|
Temperature
(°C) |
22.29
± 0.79 |
21.18
± 0.79 |
P = 0.0238* |
|
ZDV
(ml m-3) |
1.9 ±
1.7 |
1.8 ±
1.0 |
P
= 0.3867 |
|
Abundance
( |
6633 ±
7215 |
3553 ±
4188 |
P
= 0.2624 |
The
horizontal distribution of surface temperature and salinity showed temporal variability. During
these two summers, warmer temperatures (22 –
In
1994, higher salinities were observed at external sites, though in 1995, higher
values were observed at intermediate sites, in front of
In the
intermediate layers, however, the topography of 15° C isotherms suggested anticyclonic circulation near the continental slope edge (Rubín et al., 1997, 1999). This coincidence of the
subsurface circulation with the edge of the continental slope would corroborate
the notion that anticyclonic circulation in this area
seemed to be a permanent feature in summer time (Garcia et al., 2002).
Biomass
distribution
Zooplankton
displacement volume (Table 1) as calculated from the Bongo catches (250 µm)
varied between 0.5 – 6.2 ml/m3, in summer 1994 and 1.1 – 5.0 ml/m3,
in summer 1995. Distribution of biomass followed the isotherms very clearly. A
sharp decrease in ZDV was observed at the continental margin along the 200m
depth line. Very low ZDV values (0.5 – 2.0) were found in areas where
temperature in
Abundance of mesozooplankton
Mesozooplankton abundance (Table
1) was higher in 1994 (504 – 24734 ind/m3)
than 1995 (1215 – 15083 ind/m3), but the
statistical differences were not significant (p > 0.05). In the two summers
coastal-shelf tendency was observed (Figure 2). Highest numbers of organisms
were found at the shallow stations 1 and 3. The lowest mesozooplankton
abundance was found at offshore stations 9 and 10 with concentrations of 500 to
2000 ind/m3, showing densities more than
ten times less than at high density station. In 1994 the total mesozooplankton was most abundant in south area of
Guadalquivir River. However, north area between Guadiana and Guadalquivir
rivers was most abundant in
Taxonomic
composition of mesozooplankton
A
total of 15 taxa (Table 2) were identified (14 taxa in 1994 and 14 taxa in
1995). The temporal difference between sites in the number of taxa was not
significant (p > 0.05). In the two summers holoplankton
dominated the relative abundance (98%) and was mainly represented by cladocerans followed by copepods and appendicularians.
These three groups altogether made up 96.6% in 1994 and 93.9% in 1995 of the
zooplankton abundance (Table 2). Meroplankton with 2% was mainly constituted by larval
stages of decapods and barnacles. The relative importance of different taxa
varied between sites, although cladocerans were
generally dominant, with a total relative mean abundance of 75%. Copepods
constituted 17% of the total zooplankton abundance. In the NW of the Alborán Sea, Copepoda and Cladocera were the most abundant group in spring, autumn
and winter, while in summer cladocerans were the
dominant group followed by copepods and apendicularians
(Rodrígues et al., 1982; Rodríguez, 1983;
Seguin et al., 1994; Souza et al., 2005). Four species and one
genus of Cladocera were identified in decreasing
order of abundance: Penilia avirostris, Evadne spinifera, Evadne tergestina, Evadne nordmanni, and Podon
spp. Similar cladocerans’ composition was observed in
other regions of the Mediterranean Sea (Fernández de Puelles et al.,
2004; Rodrigues, 1983; Souza et al.,
2005; Zagami et
al., 1996).
|
Table 2. List of mesozooplankton taxa identified, their order, relative
abundance (A%) and frequency of occurrence (F%) in the Gulf of Cádiz,
Spain. |
|||||||
|
|
|
1994 |
|
|
|
1995 |
|
|
Taxa |
Order |
A% |
F% |
|
Order |
A% |
F% |
|
Cladocera |
1° |
86.90 |
100 |
|
1° |
65.00 |
100 |
|
Copepoda |
2° |
7.90 |
100 |
|
2° |
25.40 |
100 |
|
Appendicularia |
3° |
1.80 |
100 |
|
3° |
3.50 |
100 |
|
Cirripedia |
4° |
0.78 |
90 |
|
|
0.00 |
0 |
|
Chaetognatha |
5° |
0.70 |
90 |
|
9 |
0.51 |
90 |
|
Decapoda |
6° |
0.58 |
100 |
|
5° |
0.85 |
100 |
|
Echinodermata |
7° |
0.54 |
100 |
|
8 |
0.59 |
90 |
|
Siphonophora |
8° |
0.26 |
100 |
|
7 |
0.65 |
100 |
|
Doliolidae |
9° |
0.21 |
80 |
|
4° |
2.00 |
100 |
|
Mollusca |
10° |
0.10 |
90 |
|
12° |
0.10 |
50 |
|
Foraminiferida |
11° |
0.10 |
80 |
|
6° |
0.75 |
100 |
|
Euphausiacea |
12° |
0.05 |
50 |
|
10° |
0.40 |
90 |
|
Polychaeta
|
13° |
0.03 |
60 |
|
13° |
0.06 |
60 |
|
Hydromedusae |
14° |
0.02 |
70 |
|
11° |
0.21 |
90 |
|
Ostracoda |
|
0.00 |
00 |
|
14° |
0.02 |
30 |
A
relatively high degree of heterogeneity in zooplankton composition was found.
The result of the MRPP analysis showed a significant difference (p=0.0001),
between two summers demonstrating an elevated temporal variability in mesozooplankton community composition (Table 2).
Individual species
distribution
Cladocera showed temporal differences in their horizontal
distribution (Figure 3). During 1994 they occurred in high abundance (average =
5766 ind/m3) in a coastal and continental
shelf sites at the north and south of Guadalquivir River. In 1995 their
abundance in shelf sites decreased (average = 2310 ind/m3).
In the Gulf of Cádiz and Alborán sea (Souza et al., 2005), the dominance of cladocerans in summer was basically due to the high
abundance of Penilia avirostris,
a biological indicator of warmer waters. In the Gulf of Naples, in the inshore
station (0-50m layer), zooplankters forming the summer peak were mainly
composed of cladocerans (Mazzocchi
and Ribera d´Alcala, 1995). The importance of cladocerans in summer is a typical pattern of Mediterranean
waters (Della Croce and Bettain, 1965; Thiriot, 1972; Siokou-Frangou,
1996; Calbert et
al., 2001; Ribera d´Alcala et al., 2004. A relatively high degree of heterogeneity in
zooplankton composition was found. The result of the MRPP analysis showed a
significant difference (p=0.0001), between two summers demonstrating an
elevated temporal variability in mesozooplankton
community composition (Table 2).
Individual species
distribution
Cladocera showed temporal differences in their horizontal
distribution (Figure 3). During 1994 they occurred in high abundance (average =
5766 ind/m3) in a coastal and continental
shelf sites at the north and south of Guadalquivir River. In 1995 their
abundance in shelf sites decreased (average = 2310 ind/m3).
In the Gulf of Cádiz and Alborán sea (Souza et al., 2005), the dominance of cladocerans in summer was basically due to the high
abundance of Penilia avirostris,
a biological indicator of warmer waters. In the Gulf of Naples, in the inshore
station (0-50m layer), zooplankters forming the summer peak were mainly
composed of cladocerans (Mazzocchi
and Ribera d´Alcala, 1995). The importance of cladocerans in summer is a typical pattern of Mediterranean
waters (Della Croce and Bettain, 1965; Thiriot, 1972; Siokou-Frangou,
1996; Calbert et
al., 2001; Ribera d´Alcala et al., 2004).

The distribution of copepods was very similar in
both summers. Copepods (Figure 3) increased their abundance from 1994 (average = 526 ind/m3) to
1995 (average = 901 ind/m3) in
the north sites (p = 0.0383)
where the influence of Atlantic water is higher. The higher abundances of Appendicularia
“shift” from south area (1994) to north area (1995), but abundance was the same between two summers
(Figure 3). Appendicularia showed a smooth
coastal-shelf decrease in density with a few spots on the shelf with
irregularly high abundance.
In
Mallorca Channel (Western Mediterranean) Copepoda and
Appendicularia were the most abundant taxa (Fernández de Puelles et al. 2003, 2004). However, in the
Biscay Bay (Cantabric Sea) copepods dominate mesozooplankton abundance (Villate
and Valencia, 1997).
Other holoplanktonic taxa, such as Chaetognatha
(Sagitta spp) and meroplankton, such as Decapoda
larvae, did not show any marked temporal differences in their abundance and
horizontal distribution (Figures 3 and 4). During 1995, Doliolidae,
Euphausiacea, Siphonophora,
Foraminiferida and Hydrozoa
(Figure 4 and 5), enlarged their distribution and abundance in coastal and
shelf sites. In 1995, Echinodermata, Mollusca and Polychaeta
(Figure 5), showed the same distribution pattern and decreased their abundance
in south coastal and shelf sites. Cirripedia larvae
were present only in 1994, with most abundance in the coastal sites, but Ostracoda were present only in 1995 at the north area
(Figure 5). In the Guadiana estuary, decapod larvae were among the most
abundant taxa (Esteves et al., 2000). In the Mondego estuary the
occurrence of larval stages of benthic invertebrates, such as decapod larvae,
was mainly restricted to the summer months (Marques et al., 2006) where this pattern is related to the release of
larvae into the water column during warmer months, when the environmental
conditions are favourable (Gonçalves et al., 2003).


Correlations
between the abundance of mesozooplankton and
oceanographic variables
Abundance of copepods and appendicularians did not show a significant relationship
with oceanographic variables (temperature, salinity, ZDV and depth) (Multiple
Regression Analysis, give the results). Although abundance of the total mesozooplankton and cladocerans
in 1994 (p < 0.0005) and 1995 (p < 0.0001) showed a positive correlation
with temperature and biomass but showed a negative correlation with salinity
and depth. In the Alborán Sea the density of copepods
decreased while cladocerans showed a positive
correlation with temperature (Souza et al.,
2005). In the Mallorca channel the high zooplankton abundance, mainly due to
copepods, was found where the coolest and more saline waters were observed, and
the lowest abundance, mainly represented by siphonophores,
chaetognaths and doliolids,
was in the warmer and less saline waters, indicating the input of Atlantic
waters (Fernández
de Puelles et
al., 2004). In the coastal zone of other Mediterranean seas the differences
in zooplankton abundance are attributed to changes in temperature regime (Kovalev et al.,
2003).
ACKNOWLEDGEMENTS
The
authors would like to thank the crew of R/V Francisco de Paula Navarro and many scientists who assisted in
collecting samples at the sea and sorted the mesozooplankton.
This study was partly supported by CAPES (Ministry of Education, Brazil) as
part of the post doctoral grant (BEX 0762-03-2).
Financial support was received from the Instituto Español de Oceanografia (IEO)
within the framework of the “Ictio.Alboran-Cádiz” project.
LITERATURE CITED
Abrantes, F. 1990. The influence of
the Guadalquivir River on modern surface sediments diatom assemblages: Gulf of
Cádiz. Comun. Serv.
Geol. Portugal
76: 23-31.
Ahlstrom, E. H. and J. R. Thrailkill. 1963.
Plankton volume loss with time of preservation. Rep. Calif. Coop. Oceanic Fish.
Invest. 9: 57-73.
Calbert, A.; E. Garrido;
M. Saiz; M. Alcaraz and C.
M. Duarte. 2001. Annual zooplankton sucession in coastal
NW Mediterranean waters: the importance of the smaller size fractions. J. Plankton
Res. 23: 319-331.
Caminas, J. A. 1983. Datos preliminares sobre la biomasa zooplanctónica en el sector norrocidental
del Mar de Alboran. Boletín del Instituto Español de
Oceanografía, 1(1): 1-18.
Catalán, I. A.; J. P. Rubin; G. Navarro and L. Prieto. 2006. Larval fish distribution
in two different hydrographics situations in the Gulf
of Cádiz. Deep Sea Research II 53:
1377-1390.
Della Croce, N. e S. Bettanin. 1965. Osservazioni sul ciclo biologico di Penilia avirostris Dana del Golfo di Napoli.
Boll. Mus. Ist. Biol. Univ.
Geneova 33: 49-68.
Drake, P.; A. M. Arias;
F. Baldó; J. A. Cuesta; A. Rodríguez; A. Silva García; I. Sobrino; D. García
González and C. Fernández Delgado. 2002. Spatial and temporal variation of the nekton
and hyperbenthos from a temperate European estuary
with a regulated freshwater inflow. Estuaries 25, 451–468.
Esteves, E., T. Pina and M. A. Chícharo. 2000. The
distribution of estuaries fish larvae: Nutritional condition and co-occurrence
with predators and prey. Acta Oecol. 21: 161–173.
Fernández de Puelles, M. L; J. Valencia; J. Jansá
and A. Morillas. 2004. Hydrographical characteristics and zooplankton distribution in the
Mallorca channel (Western Mediterranean): spring 2001. ICES, Journal of Marine
Science 61: 654-666.
Fernández de Puelles, M. L.; J. M. Pinot and J. Valencia. 2003. Seasonal and interannual variability of zooplankton community in waters
off Mallorca island (Balearic Sea, Western Mediterranean): 1994-1999. Oceanol. Acta 26: 673–686.
Folkard, A. M.; P. Davies;
A. F. G. Fiúza and I. Ambar. 1997.
Remotely sensed sea surface thermal patterns in the Gulf of Cádiz and the
Strait of Gibraltar: variability, correlations, and relationships with the
surface wind field. Journal of Geophysics Research 102: 5669-5683.
García, A. M. A. y P. D. Moyano. 1991. Estados juveniles de la ictiofauna en los caños de las salinas de
García, C. M.; L. Prieto; M. Vargas; F. Echevarría; J. García-Lafuente; J.
Ruiz and J. P. Rubín. 2002. Hydrodynamics and
the spatial distribution of plankton and TEP in the Gulf of Cádiz (SW Iberian
Peninsula). Journal of Plankton Research 24(8): 817-833.
Gonçalves, F.; R. Ribeiro and A. M. V. M. Soares.
2003. Comparison
between two lunar situations on emission and larval transport of decapod larvae
in the Mondego estuary (Portugal). Acta Oecol. 24: 183–190.
Kovalev, A. V.; Mazzocchi, M. G.; Kideys, A. E.; Toklu, B. and Skryabin, V. A.
2003. Seasonal changes in the composition and abundance of zooplankton in the
seas of the Mediterranean Basin. Turk J Zool. 27: 205-219.
Mafalda Jr., P. and J. P. Rubín. 2006. Interannual variation
of larval fish assemblages in the Gulf of Cádiz (SW Iberain
Peninsula) in relation to summer oceanographic conditions. Brazilian Archives
of Biology and Technology, Curitiba 49(2): 287-296.
Marques, S. C.; U. M. Azeiteiro; J. C.
Marques; J. M. Neto and M. A. Pardal.
2006.
Zooplankton and ichthyoplankton communities in a
temperate estuary: spatial and temporal patterns. Journal of Plankton Research
28(3): 297-312.
Mazzocchi, M. G. and M. Ribera d´Alcala. 1995. Recurrent patterns in zooplankton structure
and succession in a variable coastal environment. ICES Journal of Marine
Science 52(3-4): 679-691.
Neumann-Leitão, S.; Gusmão,
L. M.; Silva, T.; Nascimento-Vieira, D. A. and Silva,
A. P. 1999. Mesozooplankton biomass and diversity in coastal and oceanic
waters off North-Eastern Brazil. Arch. Fish.
Mar. Res. 47(2/3): 153-165.
Prieto,
L.; C. M. Garcia; A. Corzo; J. Ruiz Segura and F.
Echevarría. 1999. Phytoplankton, bacterioplankton and nitrate reductase activity distribution in relation to physical
structure in the northern
Ribera d´Alcalá M. R.; F. Conversano; F.
Corato; P. Lisandro; O. Mangoni;
D. Marino; M. G. Mazzocchi; M. Modigh;
M. Montresor; M. Nardella;
V. Saggiomo; D. Sarno and
A. Zingone. 2004. Seasonal patterns in plankton communities
in a pluriannual time series at a costal
Rodríguez, J. 1983. Estudio de una comunidad planctónica
nerítica en el Mar de Alborán. Ciclo del zooplancton.
Boletín del Instituto Español de Oceanografía 1(1): 19-44.
Rodríguez, J.; A. García and V. Rodríguez. 1982. Zooplanktonic communities of the
divergence zone in the Northwestern Alboran Sea. Marine Ecology 3: 133-142.
Rubín,
J. P. 1992. El ictioplancton del mar de Alborán. Relación de su distribución espacio-temporal y
composición, con diferentes variables ambientales y con la distribución de los
peces adultos. Tesis Doctoral, Universidad de Málaga.
Rubín, J. P.; N. Cano; P. Arrate; J. García; J. Escánez;
M. Vargas y F. Hernández. 1997. El ictioplancton,
el mesozooplancton y el medio marino en el golfo de
Cádiz, estrecho de Gibraltar y sector noroeste del mar de Alborán,
en julio de 1994. Inf. Téc.
Inst. Esp. Oceanogr. 167: 1-48.
Rubín, J. P.; N. Cano; L. Prieto; C. García; J. Ruiz; F. Echevarría; A.
Corzo; J. A. Gálvez; F. Lozano; J. C. Alonso-Santos; J. Escánez;
A. Juárez; L. Zabala; F. Hernández; J. García; Lafuente, J. y M. Vargas. 1999.
La estructura del ecosistema pelágico en relación con las condiciones
oceanográficas y topográficas en el golfo de Cádiz, estrecho de Gibraltar y mar
de Alborán (sector noroeste), en julio de 1995. Inf. Téc. Inst. Esp. Oceanogr. 175:
1-73.
Siokou-Frangou,
Souza, C. S.; P. Mafalda Jr.; S. Sallés; T. Ramirez; D. Cortés; A. Garcia; J.
Mercado y M. V. Yanez. 2005. Tendencias estacionales y espaciales en la comunidad
mesozooplanctónica en una serie temporal plurianual
en el noroeste
Stevenson, R. E. 1977.
Taylor, L. R. 1961. Aggregation, variance and the mean. Nature 189: 732-735.
Thiriot, A. 1972. Les cladocères de
Méditerranée Occidentale. III. – Cycle et répartition à Bahyuls-sur-Mer
(Golfe de Lion). Synthèse des annés 1965-1969. Vie
Milieu 23: 243-295.
Vargas, M. J.; J.
García-Lafuente; J. Delgado and F.Criado. 2003. Seasonal and
wind-induced variability of sea surface temperature patterns in the
Villate F, and V. Valencia. 1997. Mesozooplancton community indicates climate changes in a
shelf area of the Bay of Biscay Throughout 1988 to 1990. Journal of Plankton
Research 19(11): 1617-1636.
Zagami, G.; F. Badalamenti;
L. Guglielmo and A. Manganaro.
1996.
Short-term variations of the zooplankton community near the Straits of Messina
(North-eastern
Página diseñada por Prof. Jesús Rafael Méndez Natera
TABLA DE CONTENIDO DE LA REVISTA CIENTÍFICA UDO
AGRÍCOLA


