
Revista Científica UDO Agrícola Volumen 7.
Número 1. Año 2007. Páginas: 49-57
Estudio comparativo de intercambio
gaseoso y parámetros fotosintéticos en dos tipos de hojas de frijol (Phaseolus vulgaris L.)
silvestre y domesticado
Comparative
study gas exchange and photosynthetic parameters in two leaf types of wild and
domesticated bean (Phaseolus
vulgaris L.)
Maritza LÓPEZ
HERRERA 1, Cecilia Beatriz PEÑA VALDIVIA2, Juan
Rogelio AGUIRRE RIVERA3, Carlos TREJO LÓPEZ2 y Ana Laura
LÓPEZ ESCAMILLA1
1, Cecilia Beatriz PEÑA VALDIVIA2, Juan
Rogelio AGUIRRE RIVERA3, Carlos TREJO LÓPEZ2 y Ana Laura
LÓPEZ ESCAMILLA1
1Laboratorio de Morfofisiología Vegetal, Centro de Investigaciones
Biológicas (CIB), Universidad Autónoma del Estado de Hidalgo (UAEH), Carretera
Pachuca-Tulancingo s/n, Ciudad Universitaria. México. CP 42184 2Botánica,
Instituto de Recursos Naturales (IRENAT), Colegio de Postgraduados, Carretera
México-Texcoco km 35.5, Montecillo, México. CP
56230 e 3Instituto de Investigación en Zonas Desérticas, Universidad Autónoma de
San Luis Potosí (UASLP), Altair 200, Col. del Llano.
San Luis Potosí, S.L.P. México. 78377. E-mails: maritza_lh2003@yahoo.com.mx,
maritzal@uaeh.reduaeh.mx, cecilia@colpos.mx, iizd@uaslp.mx,
catre@colpos.colpos.mx y lopeza@uaeh.edu.mx
![]() Autor
para correspondencia
Autor
para correspondencia
|
Recibido: 09/01/2007 |
Fin de primer arbitraje: 01/02/2007 |
Primera revisión
recibida: 16/05/2007 |
|
Fin de segundo arbitraje: 30/05/2007 |
Segunda revisión
recibida: 12/09/2007 |
Aceptado: 17/09/2007 |
RESUMEN
El objetivo de este
estudio fue comparar el intercambio gaseoso y algunos parámetros de la
fotosíntesis en hojas primarias y trifolioladas de plantas de frijol silvestre
y domesticado en la etapa vegetativa inicial, con la finalidad de evidenciar la
eficiencia fotosintética de las variantes silvestres. Muestras de dos
poblaciones de frijol silvestre originarias de Durango y Tlaxcala, México, y
los cultivares Amarillo y Bayo Mecentral se establecieron
en invernadero. Se evaluó la conductividad estomática, tasa de asimilación de
CO2 (PN), temperatura de la hoja, tasa transpiratoria, contenido de clorofila y algunos parámetros
de la fluorescencia. El análisis de varianza mostró interacción estadísticamente
significativa entre el tipo de hoja y la variante de frijol (frijol silvestre
de Durango y Tlaxcala, México, y los cultivares Amarillo y Bayo Mecentral) para contenido de clorofila tipo a y b
y el índice a/b, y la fluorescencia
inicial, variable y máxima de la clorofila. Se observó amplia heterogeneidad
entre y dentro de los materiales silvestres, independientemente de que la
semilla fuera del mismo origen, la heterogeneidad también se observó entre los
cultivares (Amarillo y Bayo Mecentral), es decir, se
encontró una amplia variabilidad interespecífica
(entre especies) e intraespecífica (dentro de una
misma especie). Se concluye que las hojas primarias
y el folíolo central de la primera hoja trifoliolada tanto de las variantes
silvestres como domesticadas, desarrolladas en un ambiente homogéneo, muestran
diferencias significativas diversas entre y dentro de las variantes silvestres
y entre las variantes silvestres y domesticadas que conducen a contrastes en PN.
Palabras clave: Conductancia estomática,
frijol, asimilación de CO2, fluorescencia, clorofila.
ABSTRACT
The aim of this
study was to compare the gas exchange and some parameters of photosynthesis in
primary and tripholiolate leaf of wild and
domesticated common bean (Phaseolus vulgaris
L.) during early vegetative stage, with de finality of show the photosynthetic
efficiency of wild common bean. Two wild common bean samples from Durango and
Tlaxcala, Mexico, and the cultivars Amarillo and Bayo
Mecentral were cultivated under greenhouse
conditions. Stomatal conductance, CO2
assimilation net rate, leave temperature, transpiratory rate, chlorophyll
content, and some parameters of the fluorescence of the chlorophyll were
evaluated. There were statistical interaction between the leave type and common
bean variant (wild common bean from Durango and Tlaxcala, México, and the
cultivars Amarillo and Bayo Mecentral)
for chlorophyll a and b, the a/b index, beside initial, variable and maxim chlorophyll
fluorescence. A high heterogeneity between and within wild samples,
independently of its origin was observed, the heterogeneity was observed also
among the cultivars (Amarillo and Bayo Mecentral), id est, a wide interspecific
(among species) and intraspecific (within species) variability was found. It
was concluded that primary and tripholiolate leaf in
wild and domesticated variants growing in homogeneous environment show
significant differences in physiological characters, all of them are diverse between and within wild variants and wild
and domesticates variants evaluated, and conduce to significant differences in
PN.
Key words: Stomatal
conductance, common bean, CO2
assimilation, fluorescence, chlorophyll content.
INTRODUCCIÓN
Actualmente
se acepta que el frijol que se cultiva y consume se originó como resultado del
proceso de domesticación del frijol silvestre (Singh, 1999). El conjunto de
diferencias morfológicas, fisiológicas y bioquímicas más evidentes que separan
las variantes silvestres de las domesticadas se consideran producto de la
domesticación (Gepts, 1999). En el frijol, este
proceso se evidenció primeramente en las estructuras vegetativas y
reproductoras, por ello se ha supuesto que las diferencias entre los frijoles
silvestres y domesticados son sólo de tipo morfológico; los cambios provocados
por la domesticación en cualquier otro nivel (anatómico, fisiológico o
bioquímico) aún están por reconocerse y evaluarse experimentalmente. Los
estudios preliminares de muestras silvestres de frijol asociados al proceso de
domesticación en su inicio se basaron en observaciones comparativas (Brücher, 1988; Miranda, 1967) y orientados principalmente a
indagar sobre los posibles centros de su origen y domesticación. Recientemente,
las investigaciones de carácter cuantitativo han evidenciado gran diversidad
morfológica y fenológica de las poblaciones silvestres cuando son cultivadas
(Aguirre R. et al., 2003; Bayuelo-Jiménez et al.,
1999; Berrocal et al., 2002; García et al., 1997; Peña-Valdivia y Aguirre,
2003); además, han aportado elementos que apoyan la idea de que los diversos
contrastes morfológicos y fenológicos dentro y entre poblaciones silvestres y
entre éstas y las variantes domesticadas parecen ser más notables que las
fisiológicas y bioquímicas (Peña-Valdivia y Aguirre, 2003; Peña-Valdivida et al.,
1996, 1998 y 1999). Entre las características morfo-fisiológicas menos
estudiadas en el frijol silvestre y su modificación durante el proceso de
domesticación están las relacionadas con el intercambio gaseoso (Peña-Valdivia et al., 1997).
Se
considera que a través del proceso de domesticación el germoplasma ha sufrido
una reducción de su variabilidad genética y, por tanto, el frijol silvestre
podría representar un recurso genético con potencial para el mejoramiento de
caracteres relacionados con la fotosíntesis (Gepts y Debouck, 1991; Lynch et
al., 1992). Estudios diversos han revelado tasas fotosintéticas menores de
los cultivares con respecto a sus parientes silvestres (Bayuelo-Jiménez
et al., 1997; Evans, 1994; García et
al., 1997; Lynch., 1992). Es probable que la selección de cultivares de mayor
rendimiento y hojas más grandes haya contribuido a este cambio, ya que
frecuentemente se han obtenido coeficientes de correlación negativos entre la
tasa fotosintética y el área de la hoja (Evans, 1994). Por otro lado, se han
demostrado diferencias genéticas en los caracteres relacionados con la
fotosíntesis entre poblaciones silvestres de frijol de diferente origen; así,
las poblaciones silvestres mexicanas presentan mayores tasas fotosintéticas que
las de otras regiones de América (Lynch et
al., 1992). Por lo anterior, el objetivo del presente estudio fue comparar
los parámetros del intercambio gaseoso, el contenido de clorofila y su
fluorescencia en las hojas de plantas jóvenes de frijol silvestre y
domesticado.
MATERIALES
Y MÉTODOS
Material Vegetal
Se
compararon dos poblaciones de frijol silvestre, una proveniente de plantas que formaban
parte de la vegetación natural de una región
Para
el desarrollo del presente estudio las semillas de todas las variantes se
multiplicaron durante el ciclo primavera-verano de 2001. Después de la cosecha
las semillas se almacenaron a 5 + 1º C hasta su utilización. Debido a
que las semillas de las muestras silvestres originales eran notablemente
heterogéneas en tamaño, se realizó una selección por estratificación por
tamaño, bajo el supuesto de que las semillas menores eran silvestres típicas y
las mayores silvestres atípicas (Peña-Valdivia et al., 2002). Así, para el propósito de este estudio, se realizó
una comparación de medias de Tukey con los datos de
las semillas, se excluyeron las semillas medianas y se utilizaron las pequeñas
o típicas y las grandes o atípicas de cada muestra silvestre (SDP, SDG, STP y
STG: semilla pequeña y grande de Durango y Tlaxcala, respectivamente). Las
semillas de los cultivares fueron estadísticamente similares, por lo que se
consideró un tamaño único (peso promedio de 268 + 2 y 286 + 3 mg
semilla-1 para el cv. Amarillo y Bayo Mecentral,
respectivamente) (Peña-Valdivia et al.,
2002).
Condiciones de cultivo
Las semillas se sembraron en
la primavera del 2004 en recipientes de un litro con una mezcla de tierra y
arena en una proporción v/v de 2:1 y cultivadas en un invernadero con
fotoperiodo natural y 15/27 º C de temperatura media mínima y máxima diaria.
Después de la emergencia, las plantas fueron regadas cada tercer día con agua y
se aplicó dos veces por semana un riego con solución nutritiva Hogland (Epstein, 1972), este procedimiento se mantuvo
durante todo el experimento. El crecimiento se supervisó hasta que las hojas
primarias y el folíolo central de la primera hoja trifoliolada alcanzaron su
máxima expansión (etapas V2 y V3, respectivamente, V2 se refiere al periodo
desde el desarrollo completo de las hojas primarias hasta el desarrollo de la
primera hoja trifoliada y V3 comprende desde el desarrollo completo de la
primera hoja trifoliada al desarrollo de la tercera hoja trifoliada) (van Schoonhoven y Pastor Corrales, 1987) y para reconocerla, se
midieron la longitud y el ancho de las hojas diariamente en 15 plantas desde el
inicio de su expansión. Las evaluaciones fisiológicas se realizaron cuando las
hojas de cada variante de frijol alcanzaron su expansión máxima.
Caracteres medidos
La
conductividad estomática (gS) (mmol m-2 seg-1), tasa de asimilación
neta (PN) (μmol m-2 seg-1),
concentración intercelular de CO2 (Ci) (μmol mol-1), temperatura de la hoja (ºC) y tasa de transpiración (mmol
m-2 seg-1) se determinaron con un sistema portátil y
abierto para análisis de gases en el espectro infrarrojo (CIRAS-1, PPSYSTEMS).
Las evaluaciones se realizaron a las 12:00 h, cuando hay mayor intensidad
lumínica. Para conocer la capacidad de las hojas de modificar su temperatura
respecto al ambiente, y debido a que ésta última cambia continuamente, se
obtuvo la diferencia de temperaturas DT (ºC) (temperatura de la hoja menos temperatura del
ambiente).
El contenido de clorofila a y b
(mg g-1 tejido foliar) se determinó con el método descrito por Arnon (1949). Los parámetros de fluorescencia de la
clorofila (U. R.): fluorescencia inicial (F0), fluorescencia
variable (Fv
= FM – F0), fluorescencia máxima (FM) y
el radio FV/FM, se midieron con un analizador portátil de
la eficiencia vegetal PEA (Plant Efficiency
Analyzer, Hansatech, King’s Lynn, GB); las hojas se mantuvieron en oscuridad
durante 20 min con los clips foliares del mismo aparato. Todas las evaluaciones
se realizaron cada tercer día por un período de dos semanas.
Diseño experimental
El
experimento se realizó mediante un diseño completamente al azar, con arreglo
factorial de los tratamientos y cuatro repeticiones. Los factores (y niveles)
fueron: variante de frijol (dos silvestres típicos, dos silvestres atípicos y
dos cultivares mejorados) y tipo de hoja (primaria y trifoliolada). Cada repetición estuvo constituida por 30 plantas, de
las que se utilizaron al azar 10 para las evaluaciones de las hojas primarias y
15 para el primer trifolio.
Los datos se sometieron a análisis de varianza (ANOVA),
comparación múltiple de medias (Tukey, P=0,05) y
significancia de las interacciones con la prueba LSMEANS. Los análisis se
realizaron con el programa estadístico SAS para computadora personal (SAS,
1989).
RESULTADOS Y
DISCUSIÓN
Intercambio
gaseoso
El análisis de varianza mostró para la
conductividad estomática (gs), interacción
estadísticamente significativa entre el tipo de hoja y la variante de frijol.
La gS -que es un parámetro que indica
indirectamente el nivel de apertura de los estomas en la hoja primaria mostró
similitud entre las seis variantes, pero la gS
del folíolo central de la primera hoja trifoliolada de las variantes STP y STG
fue entre 25 y 50 % menor que en las hojas primarias (Figura 1 B). La
interacción de la gs repercutió a su vez en la
transpiración y en la temperatura de la hoja, pues para ambas variables se
registró interacción significativa entre el tipo de hoja y la variante de
frijol. El foliolo central de la primera hoja trifoliolada tendió a calentarse
más que la hoja primaria (Figura
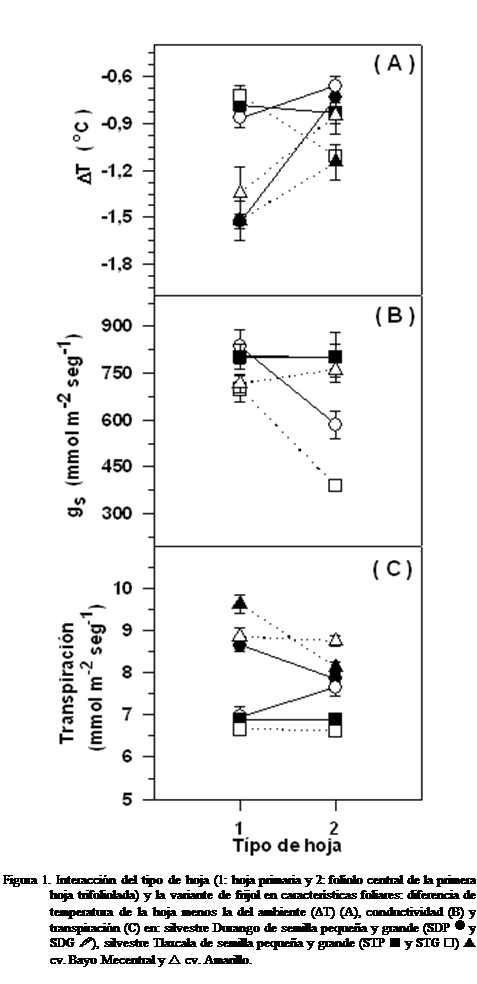
En las hojas primarias, la tasa
fotosintética (PN) es muy similar
entre las variantes estudiadas, sin embargo, se puede observar una tendencia a
que las variantes SDG y STG presenten valores un poco superiores al resto. En
el FCPHT, las variantes SDG y STG mostraron tasa fotosintética (PN)
significativamente mayor (32.0 y 11,5 %, respectivamente) que las y STP, a pesar de su procedencia respectiva
de la misma población. Las variantes de semilla pequeña presentaron una PN
estadísticamente similar a la de los cultivares (Figura
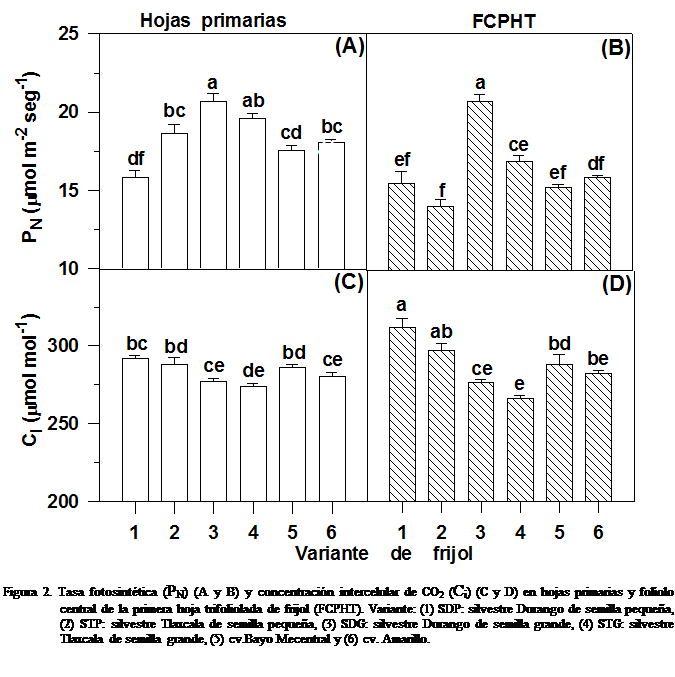
Sin embargo, la respuesta del mecanismo
estomático suele ser impredecible y frecuentemente se han observado diferencias
dentro y entre lotes de plantas y entre genotipos de una misma especie (Weyers y Meidner, 1990). En este
caso, los resultados proporcionan indicios de que dichas relaciones son
diferentes, al menos parcialmente, en algunas de las variantes silvestres, pues
aunque en proporciones bajas su Ci fue superior a lo esperado. La
experiencia indica que la respuesta estomática depende en gran medida de las
condiciones fisiológicas del material evaluado. Material experimental más
uniforme se puede obtener fácilmente de plantas de un genotipo homogéneo
desarrolladas bajo condiciones controladas. Diferencias fotosintéticas
sutiles y contrastes drásticos del rendimiento y sus componentes entre
poblaciones silvestres y domesticadas han sido documentadas por Aguirre et al. (2003), García et al. (1997) y Lynch et al. (1992). Algunos resultados del
presente estudio evidencian la posibilidad de que dichos contrastes sean
resultado parcial de los cambios en los estomas (frecuencia, tamaño,
distribución, conductividad, etc.) sucedidos durante la domesticación.
Contenido de clorofila y su fluorescencia inducida por la luz
De acuerdo con el análisis de varianza,
hubo interacción estadísticamente significativa entre el tipo de hoja y la
variante de frijol para el contenido de clorofila tipo a, tipo b y el índice a/b,
así como para los parámetros de la fluorescencia de la clorofila F0,
FV y FM (Figura 3 A-F). La representación gráfica de las
interacciones mostró mayor variación del contenido de pigmentos fotosintéticos
en las hojas primarias de los frijoles silvestres y domesticados que en el
folíolo central de la primera hoja trifoliolada (Figura
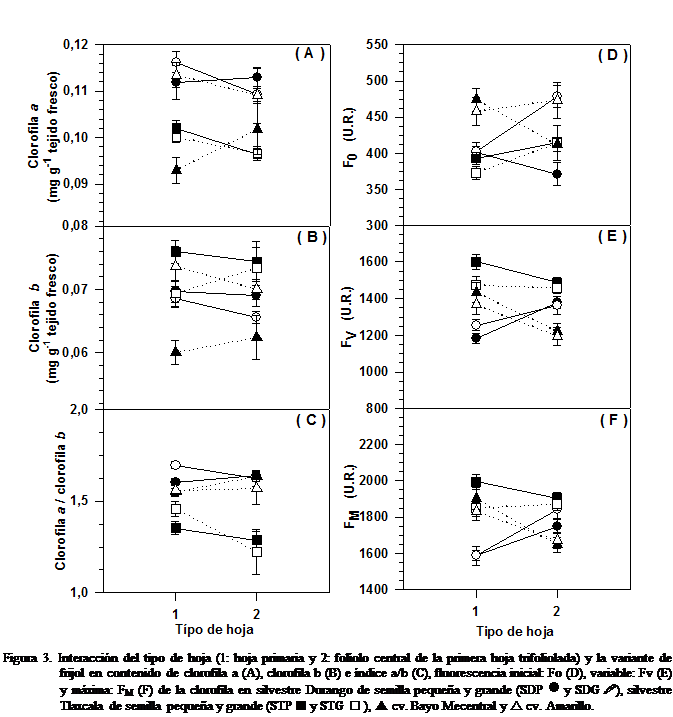
La interacción estadísticamente
significativa entre los tipos de hojas y las variantes de frijol para los
parámetros de la fluorescencia indican que el frijol modifica su respuesta
fotoquímica durante la etapa vegetativa inicial, entre V2 y V3. Así, se observa
que
En este estudio se observaron diferencias fisiológicas
relacionadas con los cambios sucedidos durante la domesticación del frijol,
hubo una mayor tasa transpiratoria y temperatura
foliar en los cultivares, las cuales podrían estar relacionadas con la menor
competencia por agua en el ambiente de cultivo; mayor eficiencia fotoquímica
del frijol silvestre, que podría estar relacionada con la respuesta para
desarrollarse con vegetación acompañante abundante; y mayor Ci en algunas variantes domesticadas lo que sugiere una
relación con la eficiencia fotosintética y fotorrespiración.
CONCLUSIONES
Las hojas primarias y el folíolo central
de la primera hoja trifoliolada tanto de las variantes silvestres como
domesticadas, desarrolladas en un ambiente homogéneo, muestran diferencias
significativas diversas entre y dentro de las variantes silvestres y entre las
variantes silvestres y domesticadas que conducen a contrastes en la tasa de asimilación
neta.
LITERATURA
CITADA
Agatti, G.; P. Mazzinghi, M. L. di
Paola, F. Fusi and G. Cecchi.
1996.
The F685/F730 chlorophyll fluorescente ratio as indicador of chilling stress in plants. Journal of Plant
Physiology. 148: 384-390.
Aguirre, R. J. R.; C. B. Peña-Valdivia and J. J. S. Bayuelo-Jiménez.
2003. Morphology, phenology and
agronomic traits of two wild Mexican common bean (Phaseolus vulgaris
L.) populations under cultivation. South African Journal of Botany 69 (3):
410-421.
Arnon, D. I. 1949. Copper enzyme in
isolated chloroplast polyphenoloxidase in Beta vulgaris. Plant Physiology 24:
1-15.
Bayuelo-Jiménez, J. S.; C. B.
Peña-Valdivia and J. R. Aguirre R. 1999. Yield components of samples of
two wild Mexican common bean (Phaseolus vulgaris
L.) populations grown under cultivation. South African Journal of Plant and
Soil 16: 197-203.
Berrocal, I. S.; J. Ortíz C. and C. B. Peña-Valdivia. 2002. Yield components, harvest index and leaf
area efficiency of a sample of wild population and a domesticated variant of
the common bean Phaseolus vulgaris. South African Journal of
Botany 68: 205-211.
Brücher, H. 1988. The wild ancestor
of Phaseolus vulgaris in South America.
In: P Gepts
(ED). Genetic Resources of Phaseolus Beans.
Kluwer Academic Publisher. Dordrecht. pp: 185-214.
Epstein, E. 1972. Mineral Nutrition of Plants: Principles and
Perspectives. John Wiley & Sons, New York, USA. 412 p.
Evans, L. T. 1994. Crop physiology: prospects for the retrospective
science. In: K. J. Boote, J. M. Bennett, T. R. Sinclair y G. M. Paulsen (EDS).
Physiology and Determination of Crop Yield. ASA, CSSA and SSSA, Madison,
Wi., USA. pp: 19-35.
García, E. 1988. Modificaciones al
Sistema de Clasificación Climática de Köppen. Instituto de Geografía, UNAM, México. 217 p.
García, H. E. R.; C.
B. Peña-Valdivia, J. R. Aguirre R. and J. S. Muruaga
M. 1997. Morphological and agronomic traits of a wild population and an
improved cultivar of common bean (Phaseolus vulgaris
L.). Annals of Botany 79: 207-213.
García, N. R.; J. R. Aguirre R., C.
Trejo, A. B. Jiménez G. and C. B. Peña-Valdivia. 2001. Contrasting effects of
shade intensity on morphology and yield components of wild and domesticated
common bean. Annual Report of the Bean Improvement Cooperative 44:204-205.
Gepts, P. and D. Debouck, 1991. Origin, domestication and evolution of
common bean (Phaseolus vulgaris L.). In: A. van Schoonhoven; O. Voysest. (Eds.)
Common Beans: Research for Crop Improvement. CAB International & CIAT,
Wallingford, UK. pp 7-53.
Lynch, J.; A. González, J. M. Tohme and J. A. García. 1992. Variation in
characters related to leaf photosynthesis in wild bean populations. Crop Science 32:
633-640.
Mckieman, M. and N. R. Baker. 1991.
Adaptation to shade of light-harvesting apparatus in Silene
dioica. Plant Cell and Environment 14:205-212.
Miranda, C. S. 1967.
Origen de Phaseolus vulgaris
L. (frijol común). Agrociencia 1: 99-109.
Morison, J. L. 1987. Intercellular CO2 concentration and stomatal response to CO2. In: E. Zeiger; G. D. Farquhar y I. R. Cowan (EDS).
Stomatal Function. Stanford, University Press, Stanford
California. 210 p.
Peña-Valdivia, C. B.; E. García H., J. R.
Aguirre R. and I. Bernal-Lugo. 1996. Tolerance of wild bean (Phaseolus vulgaris L.) seeds to suboptimal
storage conditions. Bean Improvement Cooperative 39:268-269.
Peña-Valdivia, C, B,; J. R. Aguirre R. and J. S. Bayuelo-Jiménez.
1997. Stomatal traits and photosynthesis in wild and
domesticated common bean populations. Annual Report of
the Bean Improvement Cooperative 40: 69-70.
Peña-Valdivia, C. B.; J. R. Aguirre R., E. R. García
H. y J. Muruaga M. 1998. Componentes del rendimiento de semilla de una población
silvestre y un cultivar de frijol (Phaseols vulgaris L.). Quaderni di Botanica Ambientale e Applicata 6: 181-187.
Peña-Valdivia, C. B.;
E. García H., I. Bernal-Lugo. and J. R.
Aguirre R. 1999. Seed quality of wild and domesticated common bean (Phaseolus vulgaris L.) after storage. Interciencia
24(1): 8-13 y 79-80.
Peña-Valdivia, C. B.;
R. García N., J. R. Aguirre R. and C. L. Trejo. 2002. The effects of high
temperature on dormancy and hypocotyl-root growth of wild common bean (Phaseolus vulgaris L.). Seed Science and
Technology 30: 231-248.
Peña-Valdivia, C. B. y J. R. Aguirre R. 2003. Experiencias sobre el cultivo
del frijol silvestre (Phaseolus vulgaris L.).
In: A. Casas y B. Rendón (EDS). Procesos de Evolución de Plantas Bajo
Domesticación en Mesoamérica. En prensa.
Raschke, K. 1976. How stomata resolve
the dilemma of opposing priorities. Philosophical Transaction of the Royal
Society, Series B 273: 551-560.
Statistical Analysis System (SAS). 1989. SAS Procedures Guide Version
6.04. SAS Institute. Cary, North Caroline, USA. 1290 p.
Sesták, Z. 1985. Chlorophylls and
carotenoids during leaf ontogeny. In:
Z. Sesták (ED).Photosynthesis during Leaf
Development. Dr. Junk, Publishers, Dordrecht. pp: 76-106.
Singh, S. P. 1999. Integrated genetic improvement. In: S. P. Singh (ED).Common Bean Improvement in the
Twenty-first Century. Kluwer, Dordrecht, The Netherlands. pp:
133-165.
Toro, O.; J. Thome and D. G. Debouck. 1990. Wild bean Phaseolus vulgaris description and distribution. IBPGR and CIAT, Cali, Colombia. 106 p.
van Schoonhoven, A. and M. A. Pastor-Corrales. 1987. Sistema
Estándar para la Evaluación de Germoplasma de Frijol. CIAT (International Center of
Tropical Agriculture), Cali, Colombia.
Weyers, J. D. B. and H. Meidner. 1990. Methods in Stomatal
Research & Technical. Longman, London. 223 p.
Página diseñada por Prof. Jesús Rafael Méndez Natera
TABLA DE CONTENIDO DE LA REVISTA CIENTÍFICA UDO
AGRÍCOLA


