
Revista Científica UDO Agrícola Volumen 7.
Número 1. Año 2007. Páginas: 221-227
Anatomía del tallo de lima Tahiti (Citrus latifolia Tanaka)
Stem anatomy of Tahitian lime (Citrus latifolia Tanaka)
Adolfo Enrique CAÑIZARES CHACÍN ![]() 1, Maria Elena SANABRIA2 y Eybar ROJAS2
1, Maria Elena SANABRIA2 y Eybar ROJAS2
1Instituto Nacional de Investigaciones
Agrícolas. CIAE Monagas. Vía Laguna Grande. San Agustín de la Pica. Monagas,
Venezuela y 2Posgrado de Agronomía. Universidad
Centroccidental Lisandro Alvarado, Barquisimeto, Venezuela.
E-mails:
acanizares2@hotmail.com, acanizares@inia.gob.ve y mesanabria@ucla.edu.ve ![]() Autor para correspondencia
Autor para correspondencia
|
Recibido: 29/09/2006 |
Fin de primer arbitraje: 30/10/2006 |
Primera revisión recibida: 30/10/2007 |
|
Fin de segundo arbitraje: 13/11/2007 |
Segunda revisión
recibida: 20/12/2007 |
Aceptado: 21/12/2007 |
RESUMEN
Se estudió la anatomía del tallo de la lima Tahití (Citrus
latifolia Tanaka), injertada sobre limón volcameriano (Citrus
volcameriano Pasq.), para ello se colectaron muestras del órgano con
crecimiento primario, intermedio y secundario, provenientes de plantas
cultivadas en el estado Yaracuy, Venezuela. Se seccionaron y fijaron en FAA por
12 h; se deshidrataron e incluyeron en
parafina, y se seccionaron utilizando un micrótomo de rotación. Las secciones
se tiñeron con safranina y luego de desparafinar se montaron en Permount. Las
observaciones se hicieron con un microscopio Olympus BX40. El tallo de lima Tahití con crecimiento primario
presenta epidermis uniestrata, glabra, con estomas y con células de paredes
delgadas. Los tejidos conductores primarios se disponen en haces colaterales
abiertos. En el crecimiento intermedio del tallo el tejido protector presentó
una estructura semejante a la descrita para el crecimiento primario, seguido
con una corteza, completamente parenquimatosa y
los tejidos conductores distribuidos en un anillo continuo que rodea la médula
en el centro de la sección. El felógeno
se originó a partir de la epidermis del tallo, constituyendo súber hacia la
periferia y felodermis hacia la corteza
creándose así una peridermis con la formación de las lenticelas en los sitios
donde se presentaron los estomas en el crecimiento primario. En los tallos con
crecimiento secundario el xilema y el floema secundario se disponen en anillos
continuos y la peridermis se engrosó considerablemente.
Palabras clave: Anatomía, Citrus, tallo, lima Tahití, Rutaceae,
ABSTRACT
In order to study the stem anatomy of Tahitian lime (Citrus
latifolia Tanaka), grafted on Volkameriano
lemon (Citrus volkameriano Pasq.), samples were collected with primary, intermediate
and secondary growth, at the Yaracuy state, Venezuela. These structures were
sectioned and fixed in FAA for 12 hours, they were dehidrated and demersed in
paraffin and mounted on block to make the cuts. These sections were dyed with
saffranin and mounted in permount for their description. The stem of Tahitian
lime presents unistrata epidermis, bald, with cells of thin walls. In the
primary growth, the conductive tissues: xylema and primary floemas are arranged
in collateral sheafs. In the intermediate growth, the conductive tissues form
continuous rings. The felogen begins to produce cork (suber) towards the
periphery of the stem and felodermis towards the bark, with this activity, the
formation of lenticels begins in the epidermis of the stem. In the secondary
growth, xilema and secondary floema are arranged in continuous rings.
Key words: Anatomy, Citrus, stem, Tahitian lime, Rutaceae
INTRODUCCION
Los cítricos son uno de los cultivos
frutales de mayor importancia económica en el mundo, por cuanto su
productividad se mide en términos del número de frutos cosechados por su peso, siendo el tamaño
final un parámetro de calidad para consumidores (Davies y Albrigo, 1994).
El desarrollo de la
parte aérea de los cítricos, no tiene lugar de un modo continuo, durante todo
el período de actividad vegetativa de los mismos. La aparición y desarrollo de
nuevos brotes se producen en ciclos definidos, que pueden variar de dos a cinco
anualmente. La brotación no se produce en fecha fija, varía de un año a otro y
en un mismo año, según la variedad, existiendo variaciones de carácter
individual, determinadas por las condiciones climáticas.
Flores-Vindas (1999) describió la
epidermis de los tallos de dicotiledóneas y de los cítricos con crecimiento
como uniestrata, con cutícula y con las paredes cutinizadas. En algunos casos
es posible observar abundantes tricomas y tipos tricomáticos, así como estomas.
La corteza básicamente está constituida por colénquima y parénquima, usualmente
con cloroplastos en las capas externas; y con
el segundo tejido de diferentes tipos; angular, lagunar o laminar. La
médula es casi siempre parenquimática y
puede contener cloroplastos; entre las células puede haber espacios
intercelulares conspicuos, que suelen ser más grandes hacia el centro del
órgano. Los tejidos vasculares pueden distribuirse en haces colaterales, porque
el xilema y el floema están en contacto radial, con el floema externo, o
bicolaterales con floema en ambos
extremos del haz; en los dos casos, los
haces se presentan separados por tejido
parenquimático interfascicular.
En el crecimiento secundario del tallo,
el cambium se origina de los
remanentes cambiales que quedan
en los haces caulinares y del parénquima que ocupa zonas interfasciculares. La
interpolación de tejidos vasculares secundarios entre el floema y el
xilema primario ocasiona cambios y ajustes estructurales en la anatomía del tallo, especialmente hacia
afuera del órgano. El xilema y floema secundarios sustituye a los primarios en
su función, quedando estos últimos comprimidos, aunque el parénquima vascular
puede quedar funcional. La epidermis puede permanecer y ajusta su circunferencia al crecimiento del tallo, no obstante es
sustituida por la peridermis (Flores-Vindas, 1999). La literatura acerca de las
características anatómicas de los órganos de las cítricas es escasa, hasta la fecha se conoce el
estudio realizado por Schneider (1968) quien le atribuyó al pedúnculo que
sostiene a los frutos, características de tallo. Este mismo autor consideró que
el tejido vascular primario del tallo en el género Citrus consiste en haces separados entre sí por
células grandes de parénquima. La diferenciación del
metaxilema y metafloema ocurre a partir de las derivadas procambiales
dispuestas radialmente a semejanza del cambium vascular. El tejido vascular
secundario es poroso, con vasos rodeados
por floema. El xilema secundario es
poroso, difuso, con vasos rodeados de
células parenquimáticas y fibras (Esau,
1976; Schneider, 1968).
La descripción del floema secundario se
hizo con base en una diferenciación entre el floema funcional o porción
conductiva de este tejido formada por tubos cribosos sin calosa depositada,
dispuesto en la parte interna de la corteza y adyacente al cambium vascular se
encontró el floema en desarrollo, cuyas
células derivan de las células madres
del cambium y se encuentran en proceso
de diferenciación. Por fuera de la banda de floema funcional se dispone una banda de floema en degeneración,
cuyos tubos cribosos presentaban abundante calosa, la cual una vez que se
deposita, hace que el tubo colapse y pase a formar parte del floema no
funcional, en el cual se observan además capas de fibras. Al comenzar el
crecimiento secundario la peridermis sustituye gradualmente a la epidermis
(Schneider, 1968).
El objetivo del trabajo fue realizar un
estudio de la anatomía de la lima Tahití (Citrus
latifolia Tanaka) injertada sobre limón Volcameriano (Citrus volkameriano) en las condiciones de Venezuela, como un
aporte al conocimiento de la especie.
MATERIALES
Y METODOS
Se colectaron segmentos de tallos de
plantas de lima Tahití (C. latifolia)
injertada sobre limón volcameriano (C. volkameriano) ubicadas en la
finca Hato Criollo, en la población de Temerla, Municipio Nirgua, Estado
Yaracuy, Venezuela. Las muestras se tomaron en tres estados de desarrollo:
crecimiento primario, en los extremos de las ramas (tercer flujo de
crecimiento); intermedio,
RESULTADOS
Y DISCUSION
Tallo con crecimiento primario
La epidermis se observó uniestrata y
glabra (Flores-Vindas, 1999); con las células de 1,0 1,75 mm de largo, dispuestas compactamente o interrumpida
por la presencia de estomas. El grosor de la pared externa con la cutícula midió
|
|
|
Figura
1. Sección transversal del tallo de Citrus
latifolia Tanaka, Rutaceae (lima Tahiti) con crecimiento primario. Fotomicrografía con detalles del corte: Epidermis (E),
corteza (C), haz vascular (hv), médula (M), parénquima interfascicular (pi). |

Tallo con crecimiento intermedio
Los tejidos conductores se
observaron formando anillos continuos y
el parénquima interfascicular no estaba presente por la formación de nuevas
células de floema y xilema. Los miembros de tubos cribosos del metafloema
presentan aproximadamente la misma forma y el mismo tamaño de las células
parénquimaticas de este tejido (de
|
|
|
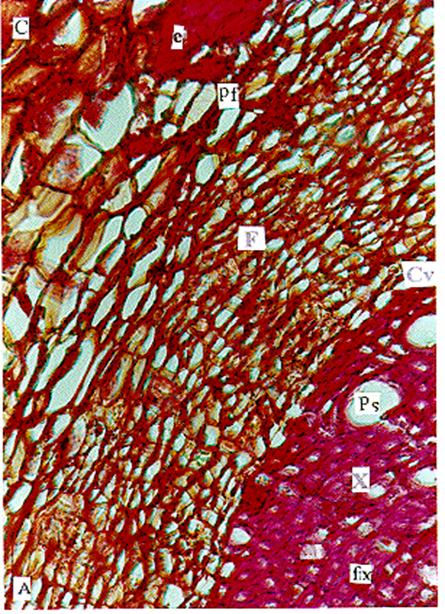
Figura 2. Sección
transversal del tallo de Citrus latifolia Tanaka, Rutaceae,
(Lima Tahití) con crecimiento intermedio. Fotomicrografía con detalles del corte. Corteza (C); esclerénquima
(e); floema (F); cambium vascular (Cv); xilema (X); fibra xilemática (fx);
poro solitario (Ps). |
Estas cavidades se forman por ruptura de
las células parenquimáticas llenas de aceites (lisis). El parénquima medular de
Los vasos del metaxilema en sección
tangencial se observaron con engrosamientos escalariformes y con las paredes terminales transversas y
perforadas. Los radios xilemáticos se presentaron homogéneos, por estar constituidos por un
sólo tipo de células (parenquimáticas)
dispuestas en uno o dos series (uni o biseriados) y dentro de las cuales se
observaron cristales semejantes a los descritos para el parénquima cortical.
Estos radios alternan con fibras xilares y los miembros de vasos del metaxilema
(Figura 3).
|
|
|
|
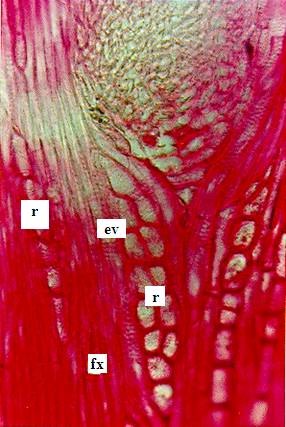
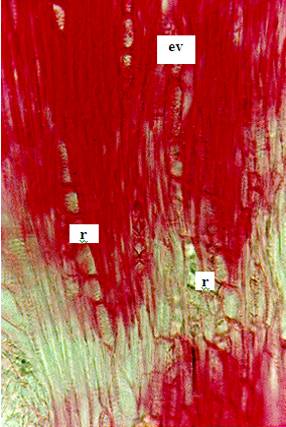
Figura 3. Sección tangencial del tallo de Citrus
latifolia Tanaka, Rutaceae, (Lima Tahití) con crecimiento intermedio. Fotomicrografía con detalles del corte: Elemento de vaso
(ev); radio xilema (r); cristal (cr); fibras xilares (fx). |
|
Tallo con crecimiento secundario
La
formación del crecimiento secundario ocasionó que las células de la corteza externa degeneraran
o se presentaran apretadas contra la periferia del tallo. La cutícula y la epidermis
fueron reemplazados, una vez iniciada la actividad del felógeno, el cual se
originó a partir de las células más externas de la corteza (primera o segunda
capa de colénquima lagunar). El felógeno comenzó a producir corcho (súber)
hacia la periferia del tallo y felodermis hacia la corteza, con esta actividad
se formó la peridermis del tallo, y se
formaron lenticelas donde se encontraban los estomas. Gradualmente a medida que
avanza el crecimiento secundario la peridermis reemplazó a la epidermis. El
xilema y el floema secundarios se disponen en anillos continuos, la médula se
redujo considerablemente. Durante la transición entre el crecimiento primario y
secundario del tallo se desarrollaron fibras a partir del parénquima
xilemático, por lo que hubo un predominio de estas células en el secundario. No
fue fácil observar exactamente el inicio de la formación del xilema secundario,
el primario si se observó claramente delimitado por células parenquimáticas del
floema secundario (Figuras 4 y 5).
|
|
|
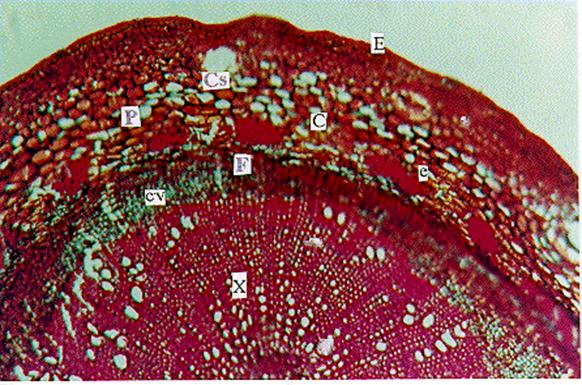
Figura 4. Sección transversal del tallo de
Lima Tahití (Citrus latifolia Tanaka) con crecimiento secundario. Fotomicrografía con detalles del corte. (E) epidermis; (C)
corteza; (P) parénquima; (Cs) cavidad secretora de aceite; (e) esclerénquima
(fibras en casquetes); (Cv) cambium vascular; (F) floema; (X) xilema. |
|
|
|
|
Figura 5. Sección
transversal del tallo de Citrus
latifolia Tanaka, Rutaceae, (Lima Tahití) con crecimiento secundario. Fotomicrografía
con detalles del corte. Epidermis (E); estomas (Est); cutícula (cu); corteza
externa (Ce); felógeno (Fe); lenticela (L); felodermis (fel), cavidad
secretora de aceites (Cs); células de relleno (cr); corteza (C); parénquima
(P); floema (F) y xilema (X). |
|


LITERATURA
CITADA
Cañizares, A. 1997. Efecto de la
cianamida de hidrógeno y el ácido e- cloroetil fosfónico sobre la defoliación,
refoliación, floración y fructificación de la lima Tahití (Citrus latifolia
Tanaka). Tesis de Grado. Magíster
Scienterum. Postgrados de Agronomía.
Universidad Centroccidental “Lisandro Alvarado”. Barquisimeto, Venezuela. p 54.
Davies, P. y L. Albrigo. 1994. Citrus, CAB Internacional Wallingford, UK.
Esau, K. 1976.Anatomía Vegetal. Ediciones Omega, Barcelona.
Flores Vindas, U. 1999. La planta estructura y función. Libro
Universitario Regional (LUR).Costa Rica.
Schneider, H. 1968. The anatomy of Citrus.
In: Reuther, Batchelor y Webber
(eds.). The Citrus Industry. Vol. 2.
University of California, Berkeley, pp. 1-86.
Página diseñada por Prof. Jesús Rafael Méndez Natera
TABLA DE CONTENIDO DE LA REVISTA CIENTÍFICA UDO
AGRÍCOLA


