
Revista Científica UDO Agrícola Volumen 12. Número 1.
Año 2012. Páginas: 74-82
Efecto
de la calidad de luz sobre el crecimiento del corocillo
(Cyperus rotundus L.)
Effect
of light quality on growth of purple nutsedge (Cyperus rotundus L.)
Nohelia M. RODRIGUEZ R.1![]() y José Vicente LAZO2
y José Vicente LAZO2
1Instituto Nacional de
Investigaciones Agrícolas (INIA). CIAE Sucre-Nueva Esparta. Campo Experimental Irapa. Estado Sucre, Venezuela y 2Universidad
Central de Venezuela, Facultad de Agronomía. Instituto de Botánica Agrícola.
Maracay, estado Aragua. E-mail; nrodríguez@inia.gob.ve ![]() Autor para correspondencia
Autor para correspondencia
|
Recibido: 09/08/2011 |
Fin de primer arbitraje: 19/01/2012 |
Primera revisión recibida: 07/02/2012 |
|
Fin de segundo arbitraje: 06/03/2012 |
Segunda revisión
recibida: 22/03/2012 |
Aceptado: 23/03/2012 |
RESUMEN
El corocillo, Cyperus rotundus L., es una maleza de amplia
distribución geográfica y difícil control. El efecto de la calidad de la luz
sobre su crecimiento fue determinado para mejorar su control. Bulbos germinados
sobre bandejas en laboratorio bajo oscuridad y luz a los 15 días, con tres
hojas y 1-7 g, se llevaron al invernadero. Se usó un diseño completamente
aleatorizado. En invernadero con suelos arenoarcillosos,
tres repeticiones y tres tratamientos. Todos tuvieron una capa de malla de
invernadero blanca, dos capas de papel
celofán blanco (MB), una verde (MV) y otra rojo (MR). La temperatura media fue
28ºC (20-35ºC), la humedad relativa media 60% (20-100%) y la radiación solar
250 a 300 g.cal.cm-2 día-1. Se realizaron cinco muestreos
cada siete días de las partes aéreas y subterráneas de las plantas. Después de
28 días, la floración ocurrió en el tratamiento MB y se observó menor número de
hojas y masa seca en las partes aéreas y subterráneas en los tratamientos MR y
MV. A los 73 días, el número de hojas y área foliar fueron menores en
tratamiento MB, mientras que en los otros tratamientos no se observaron
inflorescencias ni formación de semillas fértiles pero se estimuló formación de
mayor número cormos y hojas, siendo mayor a MR. Se concluye que la calidad de
luz es importante en la reproducción y propagación del corocillo,
por lo que la aplicación de sombra de
color rojo o verde en combinación con herbicida o control biológico surge como
alternativa para el control de esta especie.
Palabras clave: corocillo, calidad, luz, cormos, hojas
ABSTRACT
The purple nutsedge, Cyperus rotundus L., is a widely spread weed and
of difficult control. The effect of light quality on its growth was determined
to improve its control. Bulbs were germinated under darkness in the laboratory
and light at 15 days, with three leaves and 1-7 g, they were carried to the
greenhouse. A completely randomized design
was used. In greenhouse
with sandy clay soils, three replications
and three treatments. All treatments had a layer of white greenhouse
mesh, two layers of white cellophane (MB), one green (MV) and one red (MR). Average temperature was 28ºC (20-35ºC), average relative humidity 60%
(20-100%) and solar radiation 250 to 300 g. cal. cm-2 day-1.
Five samplings were made each seven days from the aerial and underground
sections of the plants. After 28 days of growth, blooming took place in the
treatment MB and a smaller number of leaves and dry weight of the aerial and
underground parts were observed in the treatments MV and MR. After 73 days, the
number of leaves and foliar area were smaller in treatment MB, while in the
other treatments neither inflorescences nor fertile seeds were observed but
formation of a larger number of corms and leaves were stimulated, MR being greater. It
is concluded that light quality is important in the reproduction and
propagations of the purple nutsedge, hence the application of green or red shade in combination
with an herbicide or biological control becomes an alternative for control of
this species.
Key words: purple nutsedge,
quality light, corms, leaves.
INTRODUCCIÓN
La duración de la luz solar por día (fotoperíodo), su
intensidad y calidad son determinantes del crecimiento, reproducción y distribución
de las especies vegetales y por supuesto de las malezas. La intensidad lumínica juega un papel
importante en el establecimiento y distribución de las malezas; algunas especies se han adaptado de tal
manera que puedan desarrollarse en sitios sombreados, lo que le permite
competir ventajosamente en plantaciones; otras al contrario necesitan una mayor
luminosidad. La luz está formada por
radiaciones de diferentes longitudes de onda; el tipo y proporción de estas
radiaciones, o sea, la calidad de luz, es susceptible de variación, dependiendo
del grosor de la capa atmosférica que atraviesa, el cual a su vez depende de la
altitud y latitud de un lugar. La
calidad de la luz afecta también la distribución geográfica de las malezas, al
permitir o no el establecimiento de diferentes especies en determinados lugares
(Vega, 1987).
La sola fuente de energía para las
plantas verdes es la energía radiante del sol.
La posición visible de esta radiación, llamada luz, es la clase más
importante de radiación en la vida de las plantas. La luz afecta la germinación de determinadas
clases de semillas, la fotosíntesis, la velocidad de crecimiento de varios
órganos, etc (Fuller et al., 1974).
La
estrategia de las plantas verdes para evitar el sombreado se basa en detectar
los cambios en la distribución espectral de la radiación filtrada a través del
dosel vegetal, así como la presencia de la vegetación adyacente mediante la
percepción de la luz reflejada. Así el fitocromo regula muchos aspectos del
desarrollo vegetal y dota a la planta de la capacidad de adaptarse a
fluctuaciones en el suministro de energía radiante regulando el desarrollo y
eficiencia del aparato fotosintético (Smith y Whitelam,
1990).
Ballaré et al
(1991) y Ballaré y Casal (2000), fueron pioneros en
demostrar la importancia de la proporción entre el rojo y el rojo lejano
AL(R/RL), como componente fundamental de la sombra entre plantas vecinas. La
captación temprana de esta señal por los
entrenudos y su relación con la densidad del follaje, la cual modula la
cantidad de radiación; también demostraron que las plantas pueden detectar la
presencia de plantas vecinas mucho antes de que estén sombreadas.
Nesser et al, (1997) demostraron que la sombra
artificial en término de radiación fotosintéticamente activa (RFA), bajo un dosel de plantas de diferentes cultivos
(maíz, pimentón, batata y frijoles) limitaba la producción de cormos de Cyperus rotundus.
En un estudio más reciente Salgado et al. (2006) encontraron que la
interferencia con el corocillo redujo el crecimiento
vegetativo en maíz; pero que la interferencia del maíz sobre corocillo originó una mayor distribución de asimilados para
los cormos.
La
mayoría de los estudios de las interacciones entre crecimiento y desarrollo del
corocillo a distintas calidades de radiación se
restringen al comparar el efecto de la sombra neutra producida por saranes o
mallas de diferente espesor que modifican la irradiancia
(cantidad de radiación) sin tomar en cuenta el otro componente de la sombra
definida por las longitudes de onda del ambiente, tal como fue reportado por
Ascencio et al. (2005).
La
respuesta a la sombra es un proceso complejo que involucra tanto la cantidad
como la calidad de la radiación y debe ser considerado en esa
dimensión Lazo y Ascencio,
(2010).
El
C.
rotundus, es una planta C4 la cual necesita de luz para su
crecimiento y desarrollo, por ello se consideró necesario estudiar como objetivo de este trabajo algunos efectos
de la calidad de la luz sobre el crecimiento de esta maleza bajo tres
tratamientos: una capa de malla de invernadero blanca + dos capas de papel
celofán blanco, una capa de malla de invernadero blanca + una capa de papel
celofán verde y una capa de malla de invernadero blanca + una capa de papel
celofán rojo.
MATERIALES Y MÉTODOS
Ubicación
El presente trabajo se realizó en el
Laboratorio y Umbráculo de Fisiología
Vegetal del Departamento de Botánica Agrícola de la Facultad de Agronomía de la
Universidad Central de Venezuela en Maracay Estado Aragua. Venezuela.
Cormos 1-7 g y plantas de C. rotundus
obtenidas después de 15 días de germinación con presencia de tres hojas.
Laboratorio
Porcentaje
de germinación y brotación,
velocidad de germinación y emergencia
Se seleccionaron aproximadamente
1000 cormos, por su tamaño y peso similar para lograr uniformidad de los mismos según metodología
usada por Cárdenas, (1992) se buscaron pesos variados de 1-7 gr. colocándose 60
cormos por bandeja total 720 en doce bandejas sobre 4 papel toalla humedecido
con agua destilada encima de las bandejas, colocadas en varios sitios del
laboratorio con luz y temperatura
ambiente 27 ºC
y en la oscuridad dentro de gavetas de los escritorios con temperatura a
24 °C. El porcentaje de germinación se determinó en forma
visual. A los tres días se observó la
germinación. Este ensayo tuvo una duración de 15 días.
Diseño experimental y tratamientos:
El diseño estadístico
utilizado para determinar el porcentaje de germinación, brotación,
velocidad de germinación y emergencia de los cormos de C. rotundus L. Aleatorizado, utilizándose 12 bandejas, 60 cormos por
bandeja, 720 en total.
Invernadero
Para las plantas en invernadero el diseño estadístico fue completamente aleatorizado con tres repeticiones y tres tratamientos de calidad de
luz, 12 plantas por tratamiento total 36 plantas. Las calidades de luz a
evaluar fueron: MB (una capa de malla de invernadero blanco más dos capas de
papel celofán blanco; MR (una capa de malla de invernadero blanca más una capa
de papel celofán rojo) y MV (una capa de
malla de invernadero blanca más una capa de papel celofán verde). El ensayo
tuvo una duración de 73 días.
Establecimiento
de plantas
Una vez germinados los cormos (tres días después de la
siembra) y las plántulas emergidas
alcanzaron tres hojas se sembraron
dentro de recipientes plásticos negros con capacidad para 5 Kg en un
suelo con textura franco arenosa, contenido de fósforo alto, potasio medio,
calcio alto, porcentaje de materia orgánica medio, pH básico de 7, 8 y
conductividad eléctrica baja. El suelo se desinfectó con 40 g del insecticida Carbofurán y se cubrió durante 20 días. Al suelo utilizado
se le realizó un análisis en el
Laboratorio de -Suelos-Planta-Nutrimentos del INIA Maracay, Estado Aragua. Se
midió el flujo cuántico con el sensor LI-COR-LI- 185B dentro y fuera del
cobertizo tomándose cuatro puntos como referencia, determinándose las
condiciones ambientales durante el período experimental: radiación solar dentro
del umbráculo, temperatura máxima, temperatura mínima y humedad relativa. Se
aplicó riego cada dos días con 400ml de agua/planta. Realizándose cinco
muestreos cada siete días, para un total de 35 días cosechándose tres plantas
por tratamiento MB, MR y MV (nueve en total) para la determinación de las
variables.
Determinación de variables
Masa seca total y por órgano (raíces, cormos,
estolones, hojas, flores), área foliar total/planta; área foliar específica,
relación sistema aéreo/raíz, sistema radical y desarrollo vegetativo y
productivo, número de hojas y número de cormos/planta, inicio de floración,
longitud y ancho de hojas. Las variables relacionadas con los rendimientos se
tomaron una vez efectuadas las muestras cada siete días durante 35 días. El
material vegetal de las tres plantas por tratamiento MB, MR, MV separado en
hojas, cormos, rizomas, raíces, inflorescencia fue colocado en bolsas de papel
y secado a estufa a 70 ºC por 72 h para la
determinación de la masa seca.
Para el estudio de las
características del sistema radical se tomó una planta por tratamiento, se
realizó el muestreo en la fase vegetativa tardía. Se separó el suelo del
sistema radical sumergiendo las bolsas en un recipiente con agua
aproximadamente unas 8 a 10 horas. Una
vez efectuado el procedimiento, se separó la parte aérea de la parte radical y
esta se preservó en una solución de etanol al 70%. Para las mediciones se
siguió el procedimiento seguido por Arnaude, (1994) el cual consiste en rehidratar las raíces
utilizando una serie decreciente de alcoholes de 30%; 20% y 10% de 15 – 30
minutos cada uno y luego colocadas en agua.
Se tiñeron con Safranina al 0,5%, con el fin de hacer más fácil las
medidas de longitud.
Para medir la longitud se realizó el método de Newman (1966) modificado por Tennant
(1975) el cual consiste en contar el número de intersecciones entre las raíces
y un patrón de líneas que se toman como referencia. Cada sistema radical una vez colocado en
agua fue trasladado a un recipiente de vidrio poco profundo, conteniendo agua jabonosa
y en el fondo se había colocado una cuadrícula de 1 cm x 1 cm. Posteriormente se contaron las intersecciones
entre las raíces y las líneas horizontales y verticales usando un controlador
manual. Finalmente la longitud radical
total para cada muestra se calculó a través de la siguiente relación:
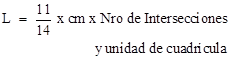
El volumen radical se
determinó utilizando el principio de desplazamiento de volúmenes. Se introdujo
la raíz en el cilindro graduado lleno de agua y se midió el volumen desplazado
por la muestra de raíces.
El diámetro promedio del
sistema radical se calculó a partir del volumen y longitud considerando la raíz
como un cilindro como lo señala Kolesnikov (1975). Posteriormente se determinó la
superficie total y densidad radical.
El área foliar se obtuvo multiplicando el largo de la hoja por el
ancho de la hoja.
Análisis estadístico
Se usó el procedimiento de Kruskal-Wallis
para pruebas no paramétricas en cada una de las variables de cinco muestreos
ejecutados cada siete días. Para evaluar la normalidad de los parámetros se uso la prueba de normalidad de Wilk-Shapiro
complementado con el método gráfico del programa Statistix
versión 4.0. Se determinaron las correlaciones de (Pearson) y la regresión
lineal de las variables muestreadas.
RESULTADOS
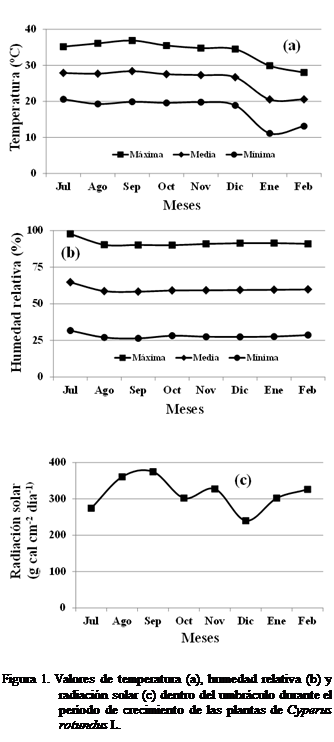
En la Figura 1 se pueden observar los
valores de temperatura (1a), humedad relativa (1b) y radiación solar (1c)
dentro y fuera del invernadero durante el tiempo de realización del
experimento. Con los valores registrados se pudo determinar que durante el
crecimiento del corocillo la temperatura máxima
alcanzada fue por encima de 35ºC, humedad relativa mayor al 80% y una radiación
solar entre 250 a 300 g cal cm-2 día-1.
Porcentaje de germinación
y brotación, velocidad de germinación y
emergencia
La germinación de los cormos etiolados colocándolos en las bandejas dentro de las
gavetas del escritorio ocurrió a los tres días, más rápidos que los expuestos a
la luz alcanzándose a obtener un 100% de cormos germinados. Por su parte los
cormos colocados en bandejas y expuestos al período de luz diaria encima de los
escritorios, tuvieron una efectividad de germinación de 70%. Los cormos
germinaron a los tres días, siendo sembrados en potes de 5 kg al tener tres
hojas formadas.
La floración ocurrió a los 28 días
después de la siembra del corocillo bajo MB.
Número de
hojas y
área foliar
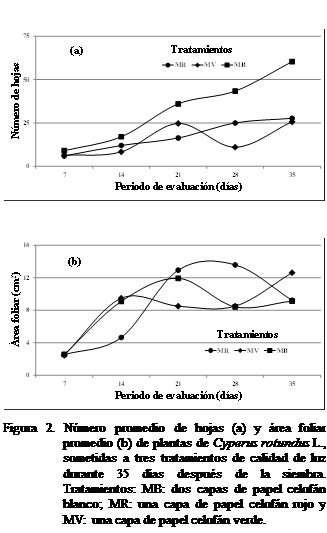
En la Figura 2a se observó el número de hojas mayor
a tratamiento MB calidad luz, desde
siete hasta 35 días después de la siembra, mayor en MV que en MR durante los
siete y 21 días. No se observaron diferencias estadísticamente significativas.
En la figura 2b el área foliar a los siete días resultó ser mayor con el
tratamiento MB , 14 y 25 días, tratamiento MV 21 y 28
MR. La variable no mostró diferencias estadísticamente significativas según Kruskal-Wallis.
Longitud de la
raíz y volumen de la raíz
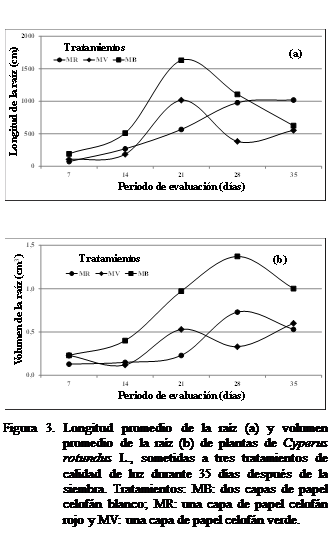
En la figura 3a se muestran los resultados de la
longitud de la raíz a los siete días después de la siembra del corocillo, observándose mayor longitud con el tratamiento
malla blanca (MB) que a malla verde (MV) y malla roja (MR). No se observaron
diferencias estadísticamente significativas según Kruskal
Wallis. Con el tratamiento MB se observó que hubo mayor crecimiento de la
longitud radicular a los 14 días pero un
comportamiento y crecimiento menor con los tratamientos MV y MR, al
comparárseles con el tratamiento MB. A los 21
días
hubo menor longitud con MV que
a MR, siendo mayor con MB. Sin embargo a
los 26
días se observó crecimiento
radicular mayor a MB. La longitud radicular obtuvo un crecimiento mayor donde
la calidad de la luz fue a MR a los 35 días. Al aplicar el tratamiento MB calidad luz, el volumen de la raíz
tuvo un comportamiento similar a MV a los siete días. Se incrementó a partir de
los 14
días después de la siembra,
hasta los 35 días (Figura 3b) Solamente se observó
diferencias estadísticamente significativas a los 21 días después de sembrado el corocillo
según Kruskal-Wallis.
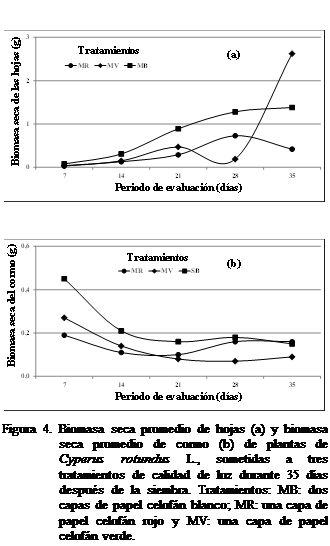
Biomasa seca de las hojas (g) y biomasa seca del cormo
(g)
La
biomasa seca de las hojas del corocillo se observó
mayor con el tratamiento MB calidad luz desde los siete días hasta 35 días después de la siembra,, seguido por
el tratamiento MV, hasta los 21 días
y MR a los 28 y 35 días (Figura 4a) La biomasa seca del cormo
a los siete y 14 días después de la
siembra del corecillo resultó mayor en MB seguido de la MV siendo menor la
biomasa alcanzada con la MR (Figura 4b) Siendo mayor a los 21 días tratamiento MR, no observándose diferencias estadísticamente
significativas. A los 28 días de haberse sembrado el C, rotundus,
la biomasa seca del cormo fue mayor a un tratamiento MR luz que a MV resultando
menor con el tratamiento MB luz. A los 35 días de siembra la biomasa seca del cormo del corocillo no tuvo variación observándose mayor en la MR que
en la MV y MB luz (Figura 4b). La
biomasa seca de las hojas mostró diferencias estadísticamente significativas,
mientras que la biomasa seca del cormo no mostró diferencias significativas
según Kruskal Wallis.
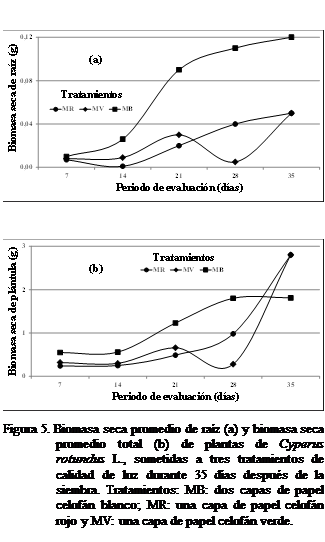
Biomasa seca (g) de la raíz (g) y biomasa seca (g)
total (g)
La
biomasa seca de la raíz del corocillo a los siete y 14 días después de la siembra se observó mayor
bajo tratamiento MB luz (Figura 5a) No se observaron diferencias
estadísticamente significativas según Kruskal-Wallis.
A los 21
días, fue mayor con el
tratamiento MB luz observándose diferencias significativas. A los 28 días después de sembrado el C. rotundus,
la biomasa seca de la raíz continuó siendo mayor con el tratamiento MB luz. No
se observaron diferencias estadísticamente significativas, mientras que a los 35 días se observó un comportamiento similar
con la MR y MV pero mayor con MB luz (Figura 5a). La mayor biomasa seca total a
los 7,
14, 21, 28 y 35 días después
de la siembra del C. rotundus, se alcanzó con el tratamiento MB luz y
la menor con el tratamiento MR, observándose a los 21 días diferencias estadísticamente significativas
(Figura 5b).
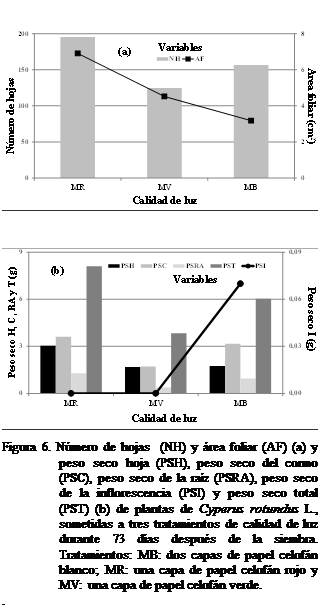
Número de hojas, longitud de hojas, ancho de
hojas, área foliar, biomasa seca de
hojas, biomasa seca del cormo, biomasa seca de la raíz, biomasa seca de la
inflorescencia y biomasa seca total
A
los 73
días después de la siembra, se
pudo observar mayor número de hojas con el tratamiento MR y menor con MV, no
observándose diferencias estadísticamente significativas según Kruskal Wallis (Figura 6a). La biomasa seca de la hoja, cormo, raíz y
total, se observaron mayor a una luz con MR, mientras que la biomasa seca de la
inflorescencia, fue menor, siendo mayor bajo tratamiento MB luz (Figura 6b). No
se observaron diferencias estadísticamente significativas.
DISCUSIÓN
El corocillo (C. rotundus)
es una de las malezas más agresivas en
los sistemas agrícolas del país y está presente en asociación con casi todos
los cultivos, siendo el control de su crecimiento una prioridad en el manejo
agronómico en condiciones de campo Lazo y Ascencio (2010). Los resultados
obtenidos con los cormos germinados en la oscuridad sobre papel toallas en
bandejas plásticas, dentro de las gavetas de los escritorios no fueron
afectados aparentemente para su brotación
coincidiendo con lo señalado por Cardenas, (1992)
cuando menciona que los tubérculos aislados en cualquier posición
independientemente de su tamaño pueden germinar y producir brotes. El
porcentaje de germinación de los cormos en bandejas encima de los escritorios
fue menor, posiblemente debido a la luz y humedad. Cárdenas, (1992) señala que
el crecimiento de las yemas, en tubérculos de corocillo
es afectado fuertemente por la luz y humedad. Durante el crecimiento del corocillo bajo tratamientos MB, MV, MR luz en el
invernadero, la temperatura máxima alcanzada, estuvo por encima de 35ºC,
humedad relativa mayor al 80% y una
radiación solar entre 250 a 300 g cal cm-2
día-1. Durante los primeros 28 días después de la siembra se observó
en el C. rotundus
una relación directa entre el número de hojas y las masas secas de las partes
aérea y subterránea cuando la intensidad de la luz fue mayor tratamiento MB.
Los resultados obtenidos coinciden con lo
señalado por Keeley y Tullen (1978) cuando mencionan
que las masas secas raíz, rizoma y cormos, generalmente aumentan con un
incremento de luz y del tiempo, señalan que la cantidad de luz tiene una
marcada influencia en el crecimiento del corocillo,
el promedio de raíces, brotes, tubérculos y masas secas aumentan en proporción
directa a la cantidad de luz .El número de hojas y las masas secas de las
partes aéreas y subterráneas fueron menores. Los resultados obtenidos en MR y
MV en relación a MB son debido tal vez a que en las plantas la luz destinada a
impulsar el proceso fotosintético es absorbida por dos tipos de pigmentos ,
clorofila y carotenoides o a la reducción de
luz la cual se encontraba influenciada por la intensidad de la luz.
Coincidiendo los resultados observados por Ramírez y Aparicio (1989), al
mencionar que tal vez podría ser debido a la falta de proceso fotosintético
ocasionado a la escasa iluminación natural con relación a las necesidades
energéticas de la planta, ya que es necesario exponer mayor cantidad de tejidos
a los factores incidentes para que pueda ocurrir una gran superficie foliar.
Giraldo y Doll (1976), mencionan, que bajo sombra C.
rotundus, produjo pocas hojas y muy delgadas, y
también se redujo la formación de rizomas y tubérculos fueron más pequeños y
producidos en menor cantidad, concordando esto con los resultados observados
durante los primeros 35 días después de sembrado el corocillo en donde el número de las hojas fue
menor y su longitud reducida en la MR bajo sombra. La floración ocurrió a los
28 días después de la siembra del corocillo bajo MB
con mayor intensidad de luz cuando el
número de hojas era mayor, Doll (1983), señala que
la floración ocurre a partir de la tercera a octava semana después de la emergencia.
A los 73 días se observó mayor número de
hojas y no hubo presencia de
inflorescencia bajo MR y MV luz., Coincidiendo con Wills (1975), quien
menciona que a menor intensidad de luz
menor número de flores. Keeley y Thullen,
(1978) señalaron que la producción de flores se redujo en ensayos efectuados
bajo sombra artificial comparándolos con los no sombreados, lo que indica que a
mayor intensidad de luz, mayor es la producción de flores A los 73 días de
sembrado el corocillo se observó un mayor número de
hojas a MR, mayor longitud y ancho de hojas que en MB, donde había mayor intensidad de luz, esto
pudiera ser explicado debido a que en MR luz estaba presente el fitocromo en su
forma activa Pfr influyendo en la morfogénesis del corocillo coincidiendo con lo mencionado por Bergarache y Moysse (1993), al
señalar que el fitocromo interacciona con los mecanismos endógenos de control
del desarrollo y provoca numerosos cambios morfológicos, fisiológicos y
bioquímicos en su forma activa Pfr . A mayor
intensidad de luz ocurre un incremento en las hojas de la planta y mayor
proceso fotosintético, ocurriendo lo mencionado por Williams (1978), cuando
indica que la fotosíntesis es un complicado proceso biológico que permite al
organismo utilizar luz visible o infrarroja, cercana como fuente de energía
metabólica, y que por lo tanto lo hace
nutrirse de compuestos minerales con bajo o nulo contenido energético.
En las plantas de C. rotundus L., MV se observó un disminución de la intensidad de luz,
modificando la calidad de luz, más que las a MR. La luz incide
directamente sobre el corocillo MB, más que a MV y MR. Fuller et al (1974),
señala que las hojas son verdes sobre todo porque la luz verde es tanto
reflejada como transmitida por la clorofila; únicamente una cantidad
relativamente pequeña de luz verde es absorbida. El manejo fisiológico de la
sombra debe ser una práctica agronómica que forma parte del control integrado
de malezas ya que tal como ha sido reportado por Mclachlan
et al. (1993), la cantidad y calidad
de radiación que se recibe debajo de un dosel a lo largo del ciclo de un
cultivo, modificando la arquitectura de las malezas es un componente
fundamental de los modelos de interferencia entre malezas y cultivos (Lazo y
Ascencio, 2010).
CONCLUSIONES
1. Los cormos colocados sobre toallas
humedecidas y etiolados germinaron en un 100%.
2. Los cormos colocados en toallas humedecidas
y sujetos a condiciones de luz, humedad y temperatura ambiente dentro del
laboratorio, germinaron aproximadamente en un 70% posiblemente debido a la luz
y a una menor humedad.
3. A mayor intensidad de luz (tratamiento
MB) estimuló en las plantas de corocillo la floración a los 28 días después e la siembra.
4. El tratamiento MR y MV inhibió la
floración en las plantas de corocillo, permitiendo
esto la no formación de semillas fértiles, como forma de reproducción y al
mismo tiempo estimuló un mayor número de cormos y hojas pudiendo ser una alternativa para su control mediante la
aplicación de un herbicida y control biológico.
LITERATURA CITADA
Arnaude,
O. 1994. Influencia del stress por
fósforo sobre el desarrollo y la respiración radical en caraota (Phaseolus vulgaris)
cv. Manaure, frijol (Vigma ungiculata) cv. Tuy, tomate (Lycopersicon
sculentum Mill.) Euphorbia heterophyla L. y Amaranthus dubius Mart. Trabajo
de grado Magíster Scientiarium en Agronomía.
Universidad Central de Venezuela. Facultad de Agronomía. Instituto de
Botánica Agrícola. Maracay. 179 pp.
Ascencio. J.; J. V. Lazo y E. Hernández.
2005. Respuesta a la calidad y cantidad de sombra en Ciperus rotundus. Revista Saber 17: 196-198.
Ballare,
C. L.; A. L. Scopel y R. A. Sanchez. 1991. Photocontrol of
stem elongation in plant neighborhoods: Effects of photon fluence
rate under natural conditions of radiation. Plant Cell Environ.14:57-65.
Ballare, C. L. y J. J. Casal. 2000 .Ligth signals perceived by crop and weed plants. Field Crop
Research 67: 149-160.
Bergareche, C. y L. Moysse. 1993. Illuminating phytochrome
functions. There is light at the end of the Tunnel. Plant Physiol
393-414.
Cárdenas, C. H. 1992. El corocillo
(Cyperus rotundus L.):
La maleza y su uso potencial. Imprenta universitaria de la Universidad Central
de Venezuela. Colección Rectorado, Facultad de Agronomía, Instituto de
Botánica, Maracay, Venezuela, 142 pp.
Doll, J. 1983. Yellow
Nudsedge Control in Field Crops. University of Wisconsin Extension Bulletin Nº 2990. 4 p.
Fuller, H. J.; Z. B. Carothers, W. W. Payne y
M. K. Balbach. 1974. Botánica.
Quinta edición. Nueva Editorial Interamericana,
S. A. de G .
V . Pag 211-212.
Giraldo, F. y J. Doll. 1976. Efecto de la
sombra sobre el crecimiento y desarrollo del coquito (Cyperus
rotundus L.) Revista COMALFI 3: 114-123
Keeley, P. y R. Thullen.
1978. Light Requirements of Yellow nutsedge
(Cyperus esculentus)
and light interceptions by crops Weed Sci. 26: 10-16
Kolesnikov, V. A. 1975. The root systems of fruit plant. Mir. Publishers,
Moscú. Traducción Ludmila
Akesno. 268 p..
Lazo, J. V.
y J. Ascencio. 2010. Efecto de diferentes calidades de luz sobre el crecimiento
del Cyperus rotundus. Bioagro 22 (2): 153-158.
Mclachlan,
S. M.; M.Tollenaar, C J. Swanton and S. F. Weise. 1993. Effect of carn induced shadin undry matter accumulation distribution, and architecture of
redroot pigweed (Amaranthus retoflecxus).Weed
Sci.41.568-573.
Nesser, C.; R. Aguero y C. J.Swanton.1997. Incident photosynthetically
active radiation as a basis for integrated management of purple nutsedge (Cyperus rotundus ).
Weed Sci. 45: 774-783.
Newman, E. L. 1966. A method of
estimating the total length of root in a sample .J. Appl. Ecol. 3, 139-145.
Ramírez, J. M. y P. J. Aparicio. 1989. Unadjustmentes
between primary and secundary electron transfer in
photosynthesis. In: Photoconversion Processess for
Energy and Chemicals. Energy
from Biomass. 5 D.O. Hall y G. Grassi (eds.). Elsevier Applied
Science, Londres.
UK.
p. 208-217.
Salgado, T.
P.; R. A. Pitelli, P. L. Alves, F. L Salvador e A .S.
Yunes. 2006. Efectos da adubacao
fosfatada nas relacoas de
interferencia inicial entre plantas de milho (Zea mays) ede tritica (Cyperus rotundus). Planta Daninha 24: 37-44.
Smith, H y G. C. Whitelan. 1990. Phytochrome a
family of photoreceptors with multiple
physiological roles. Plant Cell and Environment 13 (7): 695-707.
Tennant, D. 1975. A test of a modified line intersect method of estimating root length. J. Ecol. 63: 995-
1001.
Vega Ortega, N. 1987. Las malezas y su combate. Aspectos generales Universidad Central de
Venezuela. Ediciones de la Biblioteca. Caracas.
p. 22-23.
William, R. 1978. Photoperiod effects on the Reproductive Biology of purple nutsedge (Cyperus rotundus L. ). Weed Sci. 26: 539-592.
Wills, G. D. 1975. Effect
of light and temperature on growth of purple nutsedge.
Weed Sci
23: 93-96.
Página diseñada por Prof.
Jesús Rafael Méndez Natera
TABLA DE CONTENIDO DE LA REVISTA CIENTÍFICA
UDO AGRÍCOLA


