
Revista Científica UDO Agrícola Volumen 12. Número 1. Año
2011. Páginas: 167-173
Phenotypic and genetic parameters of
reproductive traits in Tunisian Holstein cows
Parámetros fenotípicos y genéticos de caracteres reproductivos en vacas Holstein en Túnez
Naceur M’HAMDI 1 ![]() , Rafik ALOULOU
1, Satinder K. BRAR 2, Mahdi BOUALLEGUE
3 and Mohamed Ben HAMOUDA 4
, Rafik ALOULOU
1, Satinder K. BRAR 2, Mahdi BOUALLEGUE
3 and Mohamed Ben HAMOUDA 4
1Département des Sciences Animales, Institut Supérieur Agronomique de
Chott-Mariem, BP47, 4042. Sousse, Tunisie ; 2Institut
National de la Recherche Scientifique, Quebec, Canada;
3 Faculté des sciences Mathématiques Physiques et Naturelle de
Tunis, Campus Universitaire 2092
Tunis, Tunisie and 4Institution de La Recherche et de l’Enseignement
Supérieur Agricole (IRESA), Ministère de l’Agriculture et des Ressources
Hydrauliques. Tunisie. E-mail: Email: naceur_mhamdi@yahoo.fr ![]() Corresponding author
Corresponding author
|
Received: 12/08/2009 |
First reviewing ending: 09/28/2010 |
First review received: 11/17/2010 |
Accepted: 01/04/2011 |
ABSTRACT
Various
factors influencing reproduction in dairy Holstein cows were routinely evaluated
and genetic parameters were estimated for four traits to assess fertility of
artificially inseminated cows: Calving to first service interval (CFSI),
calving interval (CI), calving to conception interval (CCI) and number of
services per conception (NSC). Data used in this investigation consisted of records
of insemination and calving events on Tunisian Holstein cows. The data included
records from 1994 to 2003 in 150 herds. The aim of the study was to investigate
the effects of non-genetic factors on reproductive traits and estimate their heritabilities. The factors examined were: month and year
of calving, herd, parity, and year-month of calving. The effect of month and
year of calving (or insemination), herd, parity and year-month of calving were
included in the model and were significant (P < 0.01) except for the number of
lactations that does not have an effect on the NSC. A decreasing efficiency in
cow fertility was observed over the last years, with a longer day for first
service interval. Heritability for fertility traits was low ranging from 0.027
for NSC to 0.067 for CI. The results suggested that more attention should be
paid to herds with too low fertility traits and that monitoring and
intervention schemes should be tested in research approaches.
Key words: fertility traits, genetic parameters, non-genetic factors, Tunisian Holstein
RESUMEN
Se evaluaron de manera rutinaria diferentes factores que influyen
en la reproducción de las vacas lecheras Holstein y se estimaron los parámetros genéticos de cuatro caracteres para evaluar la fertilidad de las vacas inseminadas
artificialmente: el intervalo
de parto a primer servicio
(IPPS), el intervalo entre partos
(IP), el intervalo de parto
a concepción (IPC) y el número
de servicios por concepción (NSC). Los datos
utilizados en esta investigación consistieron de registros de eventos de inseminación y partos en vacas Holstein de Túnez.
Los registros fueron tomados desde 1994 hasta 2003 en
150 rebaños para estudiar los efectos de factores no genéticos y estimar las heredabilidades
de los caracteres anteriormene
señalados de fertilidad. Los factores
examinados fueron: mes y año de parto, rebaño, paridad y año-mes de parto. El
efecto del mes y año de parto (o inseminación), rebaño, paridad y año-mes de
parto se incluyeron en el modelo y fueron significativos (P <0,01) con
excepción del número de lactancias que no tiene un efecto sobre el NSC. Se observe una eficiencia decreciente en la fertilidad de las vacas en los últimos años, con un día más largo para el intervalo del primer servicio. La
heredabilidad para los caracteres de la fertilidad fue baja en un
rango de 0,027 para NSC a
0,067 para el interval entre partos.
Los resultados sugieren que debería prestarse más atención a los rebaños
con rasgos muy bajos de fertilidad y que los esquemas de vigilancia e
intervención deberían ser evaluados con enfoques de investigación,
Palabras clave: Caracteres de fertilidad, parámetros genéticos,
factores no genéticos, Holstein en Túnez.
INTRODUCTION
Knowledge of reproductive performance in Tunisian dairy herds is
limited. Selection for higher yields of dairy cattle has led to a decline in
fertility due to unfavorable genetic correlations between yield and fertility
(Pryce et al., 2004). On a herd
basis, fertility in Tunisian dairy herds is not well defined and is poorly
managed. It appears difficult to correct, as the levels of the standards used
to measure many of the fertility indices are declining. Ben Hamouda
et al. (2005) on Tunisian data showed
that the reproductive performance of Holstein herds is poor. Tunisian Holstein is
the main exotic breed used for milk production in Tunisia (Ben Salem et al., 2006). During the last two
decades, the dairy sector in Tunisia went through major development programs to
increase dairy production in order to improve farmer’s lives, whose income
comes from milk sales and to ensure the national self-sufficiency in milk and
dairy products. In fact, these development programs are not in favor of
reproductive performances of Holstein which remain low.
The deterioration in fertility of dairy cows during the last two decades
currently constitutes a major problem confronted by the dairy breeders. This decline/deterioration
in reproductive performance has concerned many countries (Barbat et al.,
2005; Bosio, 2006). The results of reproductive
performances conditioned largely the breeding economic profitability and its
improvement is a part of the common imperatives for all types of production. The
actual determination of the type of traits to be included in genetic evaluation
for fertility is difficult. Earlier studies on cow reproduction possessed only
calving dates from which calving intervals or days open could be computed
assuming a standard gestation length (Jansen, 1986).
The availability of insemination data has allowed the calculation of
intervals between calving and each insemination as well as the number of
inseminations. Age at first insemination, age at conception, and the intervals
from calving to first service and first service to conception in each lactation have been important traits in several
studies (Averill et al., 2004; Jamrozik et al., 2005;
Biffani et al.,
2005). Averill et al. (2004) affirmed that reproductive performance of a cow is
an array of several traits. The heritabilities of
most reproductive traits were generally below 0.10 (Kadarmideen
et al., 2003; Wall et al., 2003).
The objectives were to identify non-genetic factors strongly associated
with reproductive performance and to estimate genetic parameters for Tunisian
Holstein female reproduction traits.
MATERIALS AND METHODS
Data Source
Data on
reproductive traits were obtained from the National Center
for Performance recording of Dairy Cattle under guardian of The Tunisian
Livestock and Pasture Office (OEP). These comprised insemination
records collected by OEP from 1994 to 2003 that were merged with pedigree,
lactation, and calving performance information to be able to calculate the
traits of interest. Pedigree records for individual cows were verified with
records from OEP. Pedigree file of all participating
animals was available and contained the ancestry of approximately 70,000
animals (about 66,000 animals and 4000 bulls). Fertility traits were
defined based on data availability in a way that would describe a complete picture
of a reproductive history for a cow. In order to describe herd fertility
retrospectively, four fertility parameters were calculated for each cow from
the insemination records: Calving to first service interval (CFSI), calving
interval (CI), Calving to conception interval (CCI) and number of services per
conception (NSC). Interval traits were expressed in days. The data file
contained herd, number of cows, date of birth, date of insemination, date of
calving, age at calving, number of lactations, number of inseminations, and
genetic group (the origin of the animal).
Data preparation
Data
consisted of recorded artificial insemination and calving events, however, data
on several variables were missing in some cases or did not correspond to the
reality. Edits performed for all traits included removal of animals having
unreasonable records; 1) with more than eight lactations; 2) which calved
before 1994 and after 2003; and 3) with incomplete records. Artificial
inseminations before 1994 were eliminated to avoid errors at the beginning of
the reproductive recording scheme. The CI that was lower than 270 days or greater
than 720 days were removed. Calving to a successful insemination and CFSI which
were less than 30 days or greater than 450 days were deleted. After editing,
records from 150 herds with 65,549 cows for the number of inseminations per
conception, calving to first service interval and calving to conception
interval, and 28,777 cows for calving interval were available for analysis.
Statistical analyses of data
The data were first
analyzed by the least squares techniques using the general linear model
procedure (SAS, 1989), to determine the effects of the various factors on
reproductive traits. CFSI, CCI and CI were analyzed using the following model.
Yijklm = μ + pi+
hj+ yk+
sl + eijklm
Where:
Yijklm = observations on the variable of interest;
μ = underlying constant,
pi = fixed effect of the ith lactation number,
hj = fixed effect of jth herd,
yk= fixed effect of kth year of calving,
sl = fixed effect of lth season of
calving,
eiklm = the random residual N (0,σ2e)
The model to analyze NSC was:
Yjklm
= μ + hj+ yk+ sl
+ ejklm
Yjklm = the observations on number of
inseminations per conception.
μ, hj, eijklm = as described in the previous model.
From the
preliminary analysis a suitable model was identified for the final estimation
of the genetic parameters. The final statistical analyses were performed with the
Derivative-Free Restricted Maximum Likelihood software (Meyer 1989) and multiple-trait animal
model to obtain variance components for CI, CFSI, CCI and NSC. The animal model
included additive genetic merit of each cow as the only random effect. To
estimate repeatability for CI, CFSI and CCI, an animal model was used to account for permanent
environmental effects common to the repeated records on the same animal.
Estimation of phenotypic, genetic and environmental trends was carried out for
CI, CFSI, CCI and NSC. The mean additive genotype in a particular year of birth
was defined as the mean predicted breeding values of cows born in that year. Consequently,
changes of mean additive genotype between the years reflected additive
genotypic differences.
The overall additive genetic trend in a trait was estimated by regressing the mean predicted breeding values on the
respective year of birth for that trait. For phenotypic trends, the adjusted
performance records were averaged within the year of birth and then regressed
on years of birth (Wakhungu, 1988; Rege and Mosi, 1989). The difference
within years between the mean predicted breeding value and the mean of the
adjusted phenotypic records reflected the component due to the non-additive
genetics and the environmental parameters.
RESULTS AND DISCUSSION
Summary of data for fertility
traits
Table 1 presents a descriptive summary of the edited data used in the
present study. The results of this study are not in agreement with the results
of Vallet et al.
(1997) who put standard objectives for CFSI of 70 days, with a percentage of
cows having CFSI > 80 days greater than 15%, CCI (90 days) with a percentage
of cows having CCI > 110 days greater than 15%, and CI (365 days) with a
percentage of cows having CI > 365 days was greater than 15%.
|
Table
1: Descriptive statistics of reproductive indices of Tunisian Holstein cattle population. |
|||||||
|
Traits |
No.
of Records |
Means
± SD |
First
quartile |
Median |
Third
Quartile |
h2 |
R |
|
NSC |
65,549 |
2.55±1.7 |
1 |
2 |
3 |
0.027 |
0.034 |
|
CFSI |
65,549 |
93.2±80.2 |
58 |
75 |
103 |
0.032 |
0.128 |
|
CCI |
65,549 |
150.9±75.7 |
84 |
138 |
194 |
0.041 |
0.135 |
|
CI |
28,777 |
444.2±101.5 |
445 |
445 |
445 |
0.063 |
0.152 |
|
NSC: Number of
services per conception; CFSI: calving to first service interval; CCI:
calving to conception interval; CI: calving intervals; h2:
Heritability; R: Repeatability and SD: Standard
Deviation. |
|||||||
Heritabilities for
the four traits are shown in Table 1; they are low and ranged from 0.027 for
NSC to 0.063 for CI. In previous studies heritability estimates were generally
weak and ranged from 0.01 to 0.05 (Maijala, 1987; Hanset et al.,
1989b) using a linear model. However, genetic improvement
of reproductive traits is very hard to achieve due to their low heritability. This
study provides genetic parameter estimates which generally confirm the
literature data: fertility traits show low heritability (<0.01) (Biffani et al.,
2005; Jamrozik et al., 2005), although some differences
do exist between traits related to time period (CI, CCI and CFSI) and score
traits (NSC). Averill et al. (2004)
reported a heritability of 0.028 for female fertility using longitudinal binary
data with Bayesian methodology. Our estimates for reproductive traits and most
estimates of other researchers are comparable and low. The reproductive trait
with the highest heritability (0.063) is CI. Considering this value and the
lower repeatability of reproductive traits (<0.03 to 0.13) (Hayes et al., 1992), it
is recognized that the reduction of one day on the delay of first service is
accompanied by an equivalent reduction of CCI (Schneider et al., 1996).
Effect of non-genetic factors on
the reproductive performance
The analysis of
variance (Table 2) showed that the number of lactations had significant effect
on CFSI, CCI and CI (P < 0.0001) with F values of 150.24, 46.90, and 5.66
respectively, but no effect was reported for NSC (P = 0.0510). Effects of herd,
calving year, calving month, and the interaction between calving year and the
month of calving and the genetic group on studied fertility traits were found
to be statistically significant (P < 0.0001). Herd had a significant
influence (P < 0.001) on traits related to time
period (CI, CCI and CFSI). Similar results have been reported in literature (Kaya,
1996; Amimo et
al., 2006).
|
Table 2. Effect estimates for selected
factors included in the final model. |
||||||||||
|
Source of |
Degree
of |
NSC |
CFSI |
CCI |
CI |
|||||
|
Variation |
Freedom |
F |
Pr>F |
F |
Pr>F |
F |
Pr>F |
F |
Pr>F |
|
|
H |
125 |
28.99 |
<0.0001 |
26.64 |
<0.0001 |
18.15 |
<0.0001 |
15.34 |
<0.0001 |
|
|
LN |
7 |
2 |
0.0510 |
149.68 |
<0.0001 |
46.36 |
<0.0001 |
5.73 |
<0.0001 |
|
|
CY |
9 |
77.79 |
<0.0001 |
83.03 |
<0.0001 |
43.70 |
<0.0001 |
74.72 |
<0.0001 |
|
|
CM |
11 |
37.72 |
<0.0001 |
28.34 |
<0.0001 |
60.98 |
<0.0001 |
7.33 |
<0.0001 |
|
|
CY*CM |
99 |
4.18 |
<0.001 |
9.92 |
<0.0001 |
12.65 |
<0.0001 |
2.29 |
<0.0001 |
|
|
GG |
16 |
5.69 |
<0.0001 |
4.87 |
<0.0001 |
8.13 |
<0.0001 |
189.61 |
<0.0001 |
|
|
Herd (H), lactation number (LN), calving year
(CY), calving month (CM) interaction calving year-calving month (CY*CM) and
genetic group (GG) |
||||||||||
The variation of CI
from one herd to another could be attributed to differences in skills of heat
detection. Effects of year of calving on CI,
CCI, CFSI and NSC were significant (P < 0.001). The year of calving is important
source of variation. Significant year of calving effects on fertility traits
have been reported in several studies (Muasya, 2005; Amimo et al., 2006).
The main effect for the model was also significant (P<0.001). CFSI, CCI and
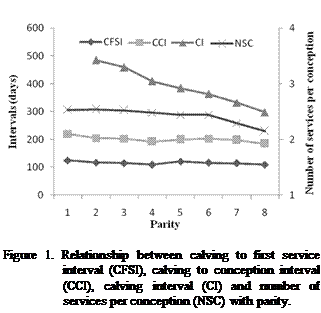
CI were found to be very long for the first lactation. The situation was
changed during the next lactations. A decrease was reported for these traits
with the lactation number between the first and the last lactations from 486 to
387 for CI, from 168 to 135 for CCI, and from 88 to 76 for CFSI (Figure 1). The
NSC was slightly decreased with the lactation number from an average value of
2.50 at first lactation to 2.11 at the eighth lactation.
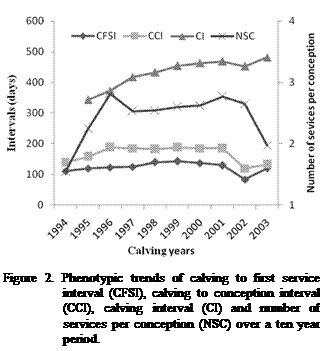
As presented in Figure 2, an elongation of CFSI, CCI and CI were observed
for many years. CI was prolonged from 13 to 17 months from 1994 to 2003,
with a noteworthy increase in 1997 (Figure 2). Fluctuation of CFSI showed that
the number of cows inseminated at less than 50 days of postpartum decreased
appreciably during the last years. It can partially explain the prolongation of
calving interval. Hence there could be a factor that contributes to larger CI.
It may be due to selection for yield or it may be due to poor management, such
as poor conception rates, poor expression or detection of oestrus
and poor nutrition. These results agree with those of Sewalem
et al., (2002) who studied reproductive
performance of Canadian dairy herds. An increase in the NSC was noticed during
the period from 1994 to 2003. The NSC ranges from 1.5 in 1994 to 2.5 at 2002
with a peak at 1996 when the NSC exceeds 2.7. A reduction has also been reported
between 2002 and 2003 in which the NSC decreased again to reach a value lower than
2. Prolongation of CFSI, CCI and CI was as a result of the important number of
belated returns (returns over 35 days exceeded 65 % of the set of returns). Chevallier and Humblo (1998) claimed
that these returns were the cause of calving interval elongation. The frequency
of longer intervals, superior in Holstein Population, suggested an elevated
embryonic mortality at a later stage (Boichard et al., 2002). Shrestha
et al. (2004) explained that higher CFSI
by the absence of heat detection at the onset of appropriate reproduction
period. On the contrary, an elongation of these intervals with lactation number
has been reported by Erb et al. (1985). Interval between calving and first insemination
decreased (Ogan, 2000) or increased (Cilek and Tekin, 2007) with
lactation number. Other authors noticed the same trend in dairy cattle compared
to beef cattle (Gregory et al., 1999b).
Fertility was known to decrease with the increase of lactation number in dairy
cattle (Weller and Ron, 1992) which was the case in this study.
Variation of fertility traits
according to calving season
The analysis of seasonal variation of the reproductive performances must be
interpreted in the light of the reciprocal influences. During the same year,
changes in management and feeding systems, temperature, humidity, and
photoperiod can be observed. The contradictory results in opposition to the
effect of season can be explained by the effect of each independent factor or
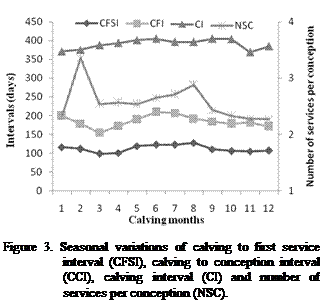
the combination of many of them at the same time. Figure 3 showed that
intervals increased during the hottest and the coldest periods of the year. An
elongation of all intervals was observed from June to August and from the first
fifteen days of November to the first fifteen days of February. Figure 3 showed
a fluctuation in the number of inseminations during the year, an increase in the
NSC during two periods; colder and hotter when NSC reached 3.5 and 3 services,
respectively. In the temperate regions, fertility is maximal in spring and
minimal in summer and winter. This was in agreement with the finding of Cilek (2009).
Westwood et al. (2002) noticed
that the anoestrus length of the post-partum was
longer when cows calved in winter, while Eldon and Olafsson
(1986) confirmed that it was shorter
for dairy cows calving in autumn. Fertility may be affected (Gregory et al., 1999b) or
may not be affected (Hageman et al.,
1991) as per seasonal variations. In tropical and
subtropical areas, many authors reported a reduction in fertility in summer
usually coinciding with the prolonged periods of elevated temperature (Weller and Ron,
1992). According to Hansen and Aréchiga
(1999), the effect of temperature on cow reproductive
performance would be translated by a decrease of heat. The modifications of the
photoperiod were not alien to the variations of reproductive performances.
Berthelot et al. (1991) mentioned the
specificities of species, mechanisms of action as well as its effects on the
puberty, calving, uterine involution and anoestrus postpartum.
CONCLUSION
Female fertility is a complex set of traits related to genetic and
environmental factors. Fertility traits showed low heritability. Reproductive
performances changed with the age of the cow often depending on previous
performances. Heritability estimates for NSC, as a trait of the cow being
inseminated was 2.7 % and was 3.2%, 4.1% and 6.3% for CFSI, CCI and CI,
respectively. There were a deterioration of the non return rates and the
elongation of the calving interval for several years. Months during the year,
lactation number, and herd affected fertility traits. In order to improve or at
least stop the deterioration trend in fertility, more emphasis on fertility
traits in selection is necessary.
ACKNOWLEDGEMENT
The authors are grateful to Hakim El Feidi, Amel Soudani and Raja Bouzidi for them assistance with grammar revision.
LITERATURE CITED
Amimo, J. O.; R. O. Mosi, J. W. Wakhungu, T. K. Muasya and B. O. Inyangala. 2006.
Phenotypic and genetic parameters of reproductive
traits for Ayrshire cattle on large-scale farms in Kenya. Livestock Research for Rural
Development. Volume
18, Article #147. Available on line:
http://www.lrrd.org/lrrd18/10/amim18147.htm.
Averill, T. A.; R. Rekaya
and K. Weigel. 2004. Genetic analysis of male and female fertility
using longitudinal binary data. J. Dairy Sci., 87: 3947-3952.
Barbat,
A. ; B. Bonaiti, F. Guillaume, T. Druet, J. J. Colleau et D. Boichard. 2005. Bilan phénotypique de la fertilité à
l'insémination artificielle dans les trois principales races laitières
françaises. Renc. Rech.
Ruminants 12 : 137-140.
Ben Hamouda, M.; M. Ben Mrad
and M. Hemdane.
2005. Genetic analyses of fertility parameters and their relations to milk
yield of Holstein-Friesian cows in Tunisia. Proceedings of
the 34th biennial session of ICAR. EAAP Publication 113: 71-76.
Ben
Salem, M.; M. Djemali, C. Kayouli and A. Majdoub. 2006. A review of environmental and management factors affecting the
reproductive performance of Holstein-Friesian dairy herds in Tunisia. Livestock Research for Rural Development. Volume
18, Article #53. Available on line: http://www.lrrd.org/lrrd18/4/sale18053.htm.
Berthelot,
X.; L. Neuhart et F. Gary.
1991. Photopériode,
mélatonine et reproduction chez la vache. Recueil de
Médecine Vétérinaire 167: 219-225.
Biffani, S. : R. Canavesi and A. B. Samore. 2005. Estimates of genetic parameters for fertility traits of Italian
Holstein-Friesian cattle. STOĈARSTVO 59 (2): 145-153.
Boichard, D.; A. Barbat
et M. Briend. 2002. Bilan phénotypique
de la fertilité chez les bovins laitiers – AERA. Reproduction, génétique et
fertilité, Paris, 6 Décembre, 5-9.
Bosio, L. 2006. Relations
entre fertilité et évolution de l’état corporel chez la vache laitière: le
point sur la bibliographie. Thèse pour obtenir le grade de
Docteur Vétérinaire; 12 Septembre 2006, Université Claude-Bernard -
Lyon I. France.
Chevallier, A. et P. Humblo. 1998. Evaluation des taux de non retour après
insémination artificielle: effet du contrôle du délai de mise à la reproduction
sur les résultats de fertilité. Renc. Rech. Ruminants 5: 75-77.
Cilek, S. and M. E. Tekin. 2007. Environmental factors affecting fertility in
cows. Indian J. Anim. Sci. 77 (3): 236-238.
Cilek, S. 2009. Reproductive traits of Holstein cows raised
at Polatli State Farm in Turley. J. Anim. Vet. Adv. 8
(1): 1-5.
Eldon, J. and T. Olafsson. 1986. The post-partum reproductive status of dairy cows in two areas in
Iceland. Acta Vet. Scand. 27: 421- 439.
Erb, H. N.; R. D. Smith, P. A. Oltenacu,
C. L. Guard, R. B. Hillman, I. P. A. Powers, M. C. Smith and M. E. White. 1985.
Path model of reproductive disorders and performance, milk fever, mastitis,
milk yield and culling in Holstein cows. J. Dairy Sci. 68 (12): 3337-3349.
Gregory,
K. E.; S. E. Echterkamp, G. E. Dickerson, L. V. Cundiff, R. M. Koch and L. D. Van Vleck.
1999. Twinning in cattle, III Effects of twinning on dystocia, reproductive
traits, calf survival, calf growth and cow productivity. J. Anim. Sci. 68:
3133-3144.
Hageman, W. H., G. E. Shook and W. J. Tyler. 1991. Reproductive performance in genetic lines selected for high or
average milk yield. J. Dairy Sci. 74: 4366- 4376.
Hansen, P. J. and C. F. Aréchiga. 1999. Strategies for managing
reproduction in the heat-stressed dairy cow. J. Dairy Sci. 82 (2): 36- 50.
Hanset, R. ; C. Michaux
and G. Detal. 1989. Genetic
analysis of some maternal reproductive traits in the Belgian Blue cattle breed.
Livest.
Prod. Sci. 23: 79- 96.
Hayes, J. F.; R. I. Cue and H. G. Monardes. 1992. Estimates of repeatability of reproductive measures in Canadian
Holsteins. J. Dairy Sci. 75: 1701-1706.
Jamrozik, J.; J. Fatehi, G. J. Kistemaker
and L. R. Schaeffer. 2005. Estimates of genetic parameters
for Canadian Holstein female reproduction traits. J. Dairy Sci. 88: 2199-2208.
Jansen,
J. 1986. Direct and maternal genetic parameters of fertility
traits in Friesian cattle. Livest. Prod. Sci. 15: 153-164.
Kadarmideen, H. N.; R. Thompson, M. P. Coffey and M. A. Kossaibati. 2003. Genetic parameters and evaluations from single- and multiple-
trait analysis of dairy cow fertility and milk production. Livest. Prod. Sci. 81: 183-195.
Kaya,
I. 1996. Parameter estimates for persistency of lactations and relationships of
persistency and milk yield in Holstein cattle. PhD Thesis, Ege University, Izmir, Turkey.
Maijala, K. 1987. Genetic control
of reproduction and lactation in ruminants. J. Anim. Breed. Genet. 104: 53-63.
Meyer, K. 1989. Restricted maximum
likelihood to estimate variance components for animal models with several
random effects using a derivative-free algorithm. Genet.
Sel. Evol. 21 (3): 317-340.
Muasya, T. K. 2005. Genetic
evaluation of the dairy cattle herd at the University of Nairobi Veterinary
Farm, MSc thesis, University of Nairobi. Kenya.
Ogan, M. 2000. The reproductive traits
of brown cow and effects of some environmental factors upon them. J. Fac.Vet. Med. Univ. Uludag 19
(3): 7-18.
Pryce, J. E.; M. D. Royal, P. C. Garnsworthy
and I. L. Mao. 2004. Fertility in
high-producing dairy cow. Livest. Prod. Sci. 86: 125- 135.
Rege, J. E. O. and R. O. Mosi. 1989. Analysis of the Kenyan Friesian breed from 1968-1984: Genetic and
environmental trends and related parameters of milk production. Bull. Anim. Hlth. Prod. Afri. 37: 267-278.
Schneider, F.; J. A. Shelford, R.
G. Peterson and L. J. Fisher. 1996. Effects
of early and late breeding of dairy cows on reproduction and production in
current and subsequent lactation. J.
Dairy Sci. 64: 1996-2002.
Sewalem, A.; G.
J. Kistemaker and B. J. Van Doormaal.
2002. Calving interval and dry period in Canadian dairy breeds. Report for
Genetic Evaluation Board.
Shrestha, H.; T. Nakao, T. Higaki, T. Suzuki
and M. Akita. 2004. Resumption of
postpartum ovarian cyclicity in high-producing
Holstein cows. Theriogenology
61(4) : 637- 649.
Statistical
Anañusis System (SAS). 1989. SAS Institute Inc.. SAS/STAT User's Guide, version 6, Fourth Edition,
Volume 2, Cary, North Carolina, United States of America.
Vallet, A.; F. Berny, J. Pimpaud, E. Lavest et L. Lagrive. 1997. Facteurs d'élevage associés à l'infécondité des troupeaux laitiers dans les Ardennes. Bulletin GTV. No 537: 23- 36.
Wakhungu, J. W. 1988.
Phenotypic, genetic and environmental trends in Kenya Sahiwal cattle. MSc. thesis,
University of Nairobi. Kenya.
Wall, E.; S. Brotherstone, J. A. Wooliams, G. Banos and M. P. Coffey. 2003. Genetic evaluation of fertility using direct and correlated
traits. J. Dairy Sci. 86: 4093-4102.
Weller, J. and M. Ron. 1992. Genetic
analysis of fertility traits in Israeli Holsteins by linear and three hold
models. J. Dairy Sci. 75: 2541-2548.
Westwood, C. T.; I. J. Lean and J. K. Garvin. 2002. Factors influencing fertility of Holstein dairy cows: A multivariate
description. J. Dairy Sci. 85(12) : 3225-3237.
Página diseñada por Prof. Jesús Rafael Méndez Natera
TABLA
DE CONTENIDO DE LA REVISTA CIENTÍFICA UDO AGRÍCOLA


